Chilesaurus
| Autor: | Mateusz Tałanda |
| Korekta: | Maciej Ziegler, Michał Siedlecki |
| Chilesaurus (czilezaur) | |
|---|---|
| Długość | do 3,5 m [1] |
| Wysokość | ok. 82 cm (w biodrach) [1] |
| Masa | ok. 100 kg [1] |
| Dieta | roślinożerny/wszystkożerny |
| Miejsce | Chile - Aysen |
| Czas | |
| Systematyka | Dinosauria |
 Rekostrukcja przyżyciowa czilezaura. Autor: Liam Elward. [1] | |
Wstęp


Chilesaurus to nieduży gatunek roślinożernego dinozaura, który żył na terenie dzisiejszego Chile około 147-148 milionów lat temu. Jego nazwa gatunkowa honoruje Diega Suáreza, który w wieku siedmiu lat znalazł pierwsze skamieniałe kości w formacji Toqui.
Materiał kopalny
Dotychczas zebrano niekompletne szkielety i pojedyncze kości 13 osobników. Mierzyły od 1,2 do 3,5 metra długości. Na ich podstawie poznano całą szyję, tułów, kończyny, część czaszki oraz przód ogona czilezaura.
Holotyp (SNGM-1935) to częściowa czaszka oraz dość kompletny szkielet pozaczaszkowy, należący do osobnika młodocianego.
Paratypy to niekompletny szkielet pozaczaszkowy też należący do osobnika młodocianego (trzy niekompletne przednie kręgi grzbietowe, częściowe żebro grzbietowe, dalsza część kości ramiennej, dalsza część k. promieniowej, niekompletna k. łokciowa, obie pierwsze, jedna druga oraz częściowa trzecia kości śródręcza, cztery paliczki palców kończyny przedniej [oba pierwsze pierwszych palców oraz pierwszy i drugi drugiego palca], niekompletne k. biodrowe, k. łonowe, k. kulszowe, k. udową, k. piszczelowe, k. skokowa, druga, trzecia i czwarta k. śródstopia oraz pierwszy i drugi paliczek drugiego palca kończyny tylnej - SNGM-1936), bardziej fragmentaryczny szkielet kończyn przednich i tylnych (k. krucze, k. ramienne [w tym jedna częściowa], k. promieniowa, k. łokciowa, pierwsza, druga i trzecia k. śródręcza, pierwszy paliczek pierwszego palca oraz pierwszy i bliższa część drugiego paliczka drugiego palca kończyny przedniej, paliczki pazurów pierwszego i drugiego palca kończyny przedniej, k. piszczelowe [w tym jedna częściowa], k. strzałkowe [w tym jedna częściowa], cztery k. śródstopia [pierwsza i druga niekompletne] oraz kilka paliczków palców kończyny tylnej [pierwszy pierwszego palca, pierwszy drugi drugiego palca, trzy pierwsze trzeciego palca, cztery czwartego palca oraz paliczki pazurów pierwszego, drugiego i trzeciego palca] - SNGM-1937) oraz kolejny fragmentaryczny szkielet pozaczaszkowy (siedem fragmentarycznych i niekompletnych kręgów grzbietowych, kilka-kilkanaście [several] żeber grzbietowych, łopatki, k. krucze, k. ramieniowe, k. promieniowe, k. łokciowe oraz obie niekompletne "ręce" składające się m.in. z pazura paliczka pierwszego palca i trzeciej k. śródręcza - SNGM-1938).
Materiał przypisany obejmuje kilka bardziej fragmentarycznych szczątków: dwie miednice (SNGM-307 i SNGM-8193), trzy trzony przednich kręgów grzbietowych (SNGM-1894, SNGM-1898 i SNGM-1903), trzon tylnego kręgu grzbietowego (SNGM-1898), niekompletną k. biodrową (SNGM-1889), bliższą część k. piszczelowej (SNGM-1895), dalszą część k. piszczelowej (SNGM-1901), trzy k. śródstopia (drugą, trzecią oraz częściową czwartą - SNGM-1890), częściową dłoń (dalszą część pierwszej k. nadgarstka, trzy k. śródręcza [pierwsza, niekompletna druga i trzecia], bliższa część pierwszego paliczka pierwszego palca, drugi paliczek drugiego palca oraz ?drugi i ?trzeci paliczek trzeciego palca - SNGM-1887) oraz dalsza część kończyny tylnej (dalsza część k. piszczelowej, dalsza część k. strzałkowej, k. skokowa, k. piętowa, dalsza część czwartej k. stępu, druga, trzecia i czwarta k. śródstopia, pierwszy i drugi paliczek drugiego palca, bliższa część pierwszego paliczka trzeciego palca, bliższa część pierwszego paliczka czwartego palca oraz niekompletny paliczek pazura - SNGM-1887).
Niezwykła budowa
Znalezione kości sugerują, że czilezaur miał małą głowę. Szczęki były krótkie, a pomarszczona powierzchnia kości przedszczękowej sugeruje istnienie keratynowej pokrywy. Czilezaur mógł mieć coś na kształt dzioba. Zęby rosły wysokie, liściokształtne i były prawie pozbawione karbowania na krawędziach. Skierowane były do przodu pyska. Niskie a długie kręgi szyjne tworzyły zgrabną szyję. Kręgi szyjne i pierwsze grzbietowe miały parę podzielonych przegrodą pleuroceli - otworów charakterystycznych dla teropodów. Czilezaur był dwunożny. Jego ręce były prawie dwa razy krótsze od nóg i kończyły się tylko dwoma palcami. Trzeci palec dłoni był zredukowany. Miednica przypomina inne teropody z jedną istotną różnicą. Kość łonowa jest skierowana do tyłu jak u dinozaurów ptasiomiedniczych, ptaków i terizinozaurów. Nie jest jednak rozszerzona na końcu jak u dwóch ostatnich grup. Przyczepy mięśni na nogach są odmienne od teropodzich, a nawiązują do zauropodomorfów i dinozaurów ptasiomiednicznych. Teropody mają pierwszy palec stopy zazwyczaj mały i skierowany do tyłu lub na bok. Tymczasem stopa czilezaura miała wszystkie cztery palce dobrze rozwinięte i skierowane do przodu - tak jak wczesne ptasiomiedniczne i zauropodomorfy oraz zaawansowane terizinozaury.
Tak niezwykła mieszanka cech skłoniła niektórych badaczy do hipotezy, że czilezaur jest w rzeczywistości prymitywnym przedstawicielem dinozaurów ptasiomiednicznych, a nie teropodem. (Baron & Barret, 2017).

Szczęki czilezaura z zachowanymi zębami. U góry kość szczękowa i przedszczękowa, na dole kość zębowa. Objaśnienia: alv - miejsce na zęby, cr - korona zęba, ro - korzeń. Źródło: Novas i in., 2015.

Kręg czilezaura z otworami (pleurocelami) charakterystycznymi dla teropodów. Objaśnienia: poz i prz - wyrostki stawowe (zygapofyzy), ppl - pleurocel. Źródło: Novas i in., 2015.
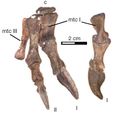
Dłoń czilezaura ze zredukowanym trzecim palcem. Objaśnienia: c - kość nadgarstka, mtc - kość śródręcza. Źródło: Novas i in., 2015.

Prawa strona miednicy czilezaura widoczna od boku. Widać jak kość łonowa jest skierowana do tyłu (w lewą stronę obrazka) jak dinozaurów ptasiomiednicznych. Wybrane objaśnienia: is - kość kulszowa, p - kość łonowa. Źródło: Novas i in., 2015.




Jednakże, Müller i współpracownicy w 2018 roku, po ponownej analizie filogenetycznej, stwierdzili, że czilezaur był członkiem kladu zauropodomorfów, będącym taksonem siostrzanym do kladu zawierającego efraazję i innych bardziej zaawansowanych zauropodomorfów. Autorzy poprzedniego artykułu (Baron & Barret, 2018) podtrzymują, iż czilezaur był przedstawicielem Ornithischia, będąc najbardziej bazalnym przedstawicielem tego kladu. Potwierdzają to również nowsze badania (np.: Müller & Dias-da-Silva, 2019 ; Müller, 2022), jednak nadal zwracają uwagę na trudności związane z ustaleniem pokrewieństw tego dinozaura. Paul (2024) zaliczył czilezaura do tetanurów, jednak zaznaczył, że jego pokrewieństwo wewnątrz tego kladu jest niedostatecznie określone. Zasugerował również pokrewieństwo czilezaura do eszanozaura. Baron (2024) stwierdził, że z powodu mozaiki cech różnych grup dinozaurów obecnej u omawianego dinozaura, czilezaur może być albo teropodem albo dinozaurem ptasiomiednicznym, przy czym jego przynależność do tej pierwszej grupy, a w jej obrębie do tetanurów jest niewiele bardziej wspierane przez różne matryce cech morfologicznych do analiz filogenetycznych niż do drugiej.
Paleobiologia

Czilezaur jest kolejnym przykładem teropoda ( o ile nim był), który dostosował się do roślinożerności. Do tej pory znano takie przypadki głównie wśród najbardziej zaawansowanych teropodów. Wyjątkiem był jurajski limuzaur. Jak się okazuje takie sytuacje nie były rzadkie także we wczesnej ich ewolucji. Ze względu na zmianę diety z mięsożernej na roślinożerną, przebudowie uległo prawie całe ciało przodków chilezaura. Zęby, z nożowatych, stały się liściokształtne. Czilezaur skubał nimi pędy roślin. By lepiej trawić pokarm, rozbudowie uległ jego układ pokarmowy. Wymusiło to przesunięcie się kości łonowych do tyłu, by brzuch stał się bardziej pojemny. Z powodu cięższego brzucha nogi stały się krótsze, ale mocniejsze. Czilezaur nie był typem szybkiego biegacza. Przez większość czasu pasł się powoli wśród roślinności. Był głównym roślinożernym konsumentem w swoim ekosystemie. Jego chód musiał bardziej przypominać dinozaury ptasiomiedniczne i prozauropody, niż dinozaury drapieżne.

![[1]](https://commons.wikimedia.org/wiki/File:Chilesasaurus.png){kind=link}
Znalezione okazy czilezaura miały wyraźnie zgięte ręce w łokciach i nadgarstkach. Takie ich ustawienie przypomina to występujące u dinozaurów bliskich ptakom, które znaleziono w pozycji spoczywającej, takich jak mei, sinornitoid i albinyk. Tak wygięty nadgarstek do dołu i na bok najwyraźniej cechuje wszystkie tetanury, a nie tylko ptaki i ich najbliższych krewnych. Natomiast ustawienie całego ciała w pozycji spoczynkowej jak u dzisiejszych ptaków znane jest także u guaibazaura, saltopa i skleromochla. Musiało być więc charakterystyczne już dla przodków dinozaurów. Jedynie nogi w szkieletach czilezaura są wyprostowane do tyłu, co różni go od "śpiących dinozaurów". Naukowcy zatem sugerują, że czilezaur był przytomny w chwili pogrzebania go przez osad. Prawdopodobnie odpoczywał lub spokojnie żerował wśród roślinności, gdy zastała go śmierć.
Molina-Pérez i Larramendi (2019) oszacowali maksymalną szybkość czilezaura na 27,6 km/h (7,4 m/s).
Czilezaur żył w wilgotnym lesie składającym się z drzew Podocarpoxylon, pokrewnych naszemu cisu, oraz paproci. Innymi mieszkańcami tego lasu były duże teropody i zauropody (np.: diplodocydy i titanozauriformy) oraz krokodyle ( w tym przedstawiciel grupy Neosuchia Burkesuchus).
Paleobiogeografia
W czasach czilezaura głównymi roślinożercami średniej wielkości były dinozaury ptasiomiedniczne takie jak kamptozaur, dysalotozaur, dryozaur, kentrozaur, czy stegozaur. Tymczasem formacja Toqui była zdominowana przez chilezaury. Jest to zaskakujące biorąc pod uwagę fakt, że w późnej jurze fauny dinozaurów były bardzo podobne do siebie na całym świecie. Czilezaur reprezentuje nową, nieznaną wcześniej linię dinozaurów, która była endemiczna dla zachodniej części Gondwany.
Finsterbusch i Moreno (2018) porównali stopę czilezaura z czteropalczastymi tropami, przyporządkowanymi do ichnogatunków Exallopus lovei ( odkryte w stanie Wyoming, w USA) oraz Picunichnus benedottei( odnalezione w prowincji Neuquén, w Argentynie) i wywnioskowali, że twórcy tych tropów mogli być spokrewnieni z tym enigmatycznym dinozaurem. jeśli wnioski te okazałyby się prawdziwe, dinozarury spokrewnione z czilezaurem mogłyby również być obecne na terenie zachodniej Laurazji. Ponadto, Lockley i współpracownicy (2018) zasugerowali, że czilezaur mógł być jednym z autorów tropów, zaliczanych do ichnorodzaju Anomoepus, co również potencjalnie poszerza jego zasięg geograficzny.
Spis gatunków
| Chilesaurus | Novas, Salgado, Suárez, Agnolín, Ezcurra, Chimento, de la Cruz, Isasi, Vargas i Rubilar-Rogers, 2015 |
| C. diegosuarezi | Novas, Salgado, Suárez, Agnolín, Ezcurra, Chimento, de la Cruz, Isasi, Vargas i Rubilar-Rogers, 2015 |
Literatura
Baron, M.G. (2024). "The effect of character and outgroup choice on the phylogenetic position of the Jurassic dinosaur Chilesaurus diegosaurezi". Palaeoworld. 33 (1): 142–151. [2]
Baron, M.G., Barrett, P.M. (2017). "A dinosaur missing-link? Chilesaurus and the early evolution of ornithischian dinosaurs". Biology Letters, 13 (8) nr art. 20170220. DOI: 10.1098/rsbl.2017.0220
Baron, M.G., Barrett, P.M. (2018). "Support for the placement of Chilesaurus within Ornithischia: a reply to Müller et al.". Biology Letters, 14 (3) [[3]]
Chimento, N.R., Agnolín, F.L., Novas, F.E., Ezcurra, M.D., Salgado, L., Isasi, M.P., Suárez, M., De La Cruz, R., Rubilar-Rogers, D., Vargas, A.O. (2017). "Forelimb posture in Chilesaurus diegosuarezi (Dinosauria, Theropoda) and its behavioral and phylogenetic implications". Ameghiniana, 55 (5): 567-5765 doi: 10.5710/AMGH.11.06.2017.3088
Fintersbush, O., Moreno, K. (2018) "Following the tracks of Chilesaurus: A new look into the record of dinosaur tetradactyl footsprints. A preliminary study" Advances en Paleontología Chilena [[4]]
Lockley, M.G., Foster, J.R., Gierlinski, G.D., Kenchiro, S. (2018). "Upper Triassic Tetrapod Tracks Assemblage From The Chinle Group, Moab Area, Utah". [W] Lucas, S.G. & Sullivan, R.M. (red) Fossil Record 6 (2) (2018), New Mexico Museum of Natural History Bulletin 79.[[ https://books.google.pl/books?hl=pl&lr=&id=ZFfPDwAAQBAJ&oi=fnd&pg=PA417&dq=chilesaurus&ots=nfQOfUGvoV&sig=Ec5d03EKMMhSDDPNNlEyjcBXUec&redir_esc=y#v=onepage&q=chilesaurus&f=false]]
Molina-Pérez, R. & larramedi, A. (2019) "Dinosaur Facts and Figures. The Theropods and Other Dinosauriformes", Princeon Univeristy Press
Müller, R.T. (2022) "The effect of character and outgroup choice on the phylogenetic position of the Jurassic dinosaur Chilesaurus diegosaurezi" Paleoworld, in press [[5]]
Müller, R.T., Dias-sa-Silva, A. (2019). "Taxon sample and character coding deeply impact unstable branches in phylogenetic trees of dinosaurs" Historical Biology 31 (8): 1089-1092. [[6]]
Müller, R. T., Pretto, F.A., Kerber, L., Silva-Neves, E. & Dias-da-Silva, S. (2018). "Comment on ‘A dinosaur missing-link? Chilesaurus and the early evolution of ornithischian dinosaurs'". Biology Letters, 12 (3)[[7]]
Novas, F.E., Salgado, L., Suárez, M., Agnolín, F.L., Ezcurra, M.D., Chimento, N.R., de la Cruz, R., Isasi, M.P., Vargas, A.O., Rubilar-Rogers, D. (2015). "An enigmatic plant-eating theropod from the Late Jurassic period of Chile". Nature, 522 : 331-334. doi:10.1038/nature14307
Salgado, L., De La Cruz, R., Suárez, M., Gasparini, Z., Fernández, M. (2008). "First Late Jurassic dinosaur bones from Chile". Journal of Vertebrate Paleontology. 28 (2): 529–534. [8]
Suárez, M., De La Cruz, R., Fanning, M., Novas, F., Salgado, L. (2016). "Tithonian age of dinosaur fossils in central Patagonian, Chile: U–Pb SHRIMP geochronology". International Journal of Earth Sciences, 105 (8): 2273-2284. DOI: 10.1007/s00531-015-1287-7
Paul, G.S. (2024) "Princeton Field Guide to Dinosaurs (3 ed.)". Princeton Univerity Press.