Spinosaurus: Różnice pomiędzy wersjami
m |
m (→Bibliografia) |
||
| (Nie pokazano 118 wersji utworzonych przez 3 użytkowników) | |||
| Linia 1: | Linia 1: | ||
| − | |||
| − | |||
{{DISPLAYTITLE:''Spinosaurus''}} | {{DISPLAYTITLE:''Spinosaurus''}} | ||
| − | + | {{Opis | |
| − | { | + | |Autor = [[Maciej Ziegler]], [[Kamil Kamiński]], [[Tomasz Sokołowski]] |
| − | | Autor | + | |Korekta = [[Tomasz Skawiński]], [[Marcin Szermański]] |
| − | + | |nazwa = ''Spinosaurus'' (spinozaur) | |
| − | + | |długość = > 11-13 m / 12-14 m | |
| − | + | |wysokość = | |
| − | [[ | + | |masa = 3,2-4,2 t / 5,4-7,1 t |
| − | + | |dieta = | |
| − | [[ | + | |miejsce = [[Egipt]]<br><small>([[formacja]] [[Bahariya]])</small> |
| − | | | ||
| − | [[Tomasz Skawiński]] | ||
| − | |||
| − | [[Marcin Szermański]] | ||
| − | | | ||
| − | </small> | ||
| − | + | [[Maroko]]<br> | |
| + | <small>[[Kem Kem]] (=[[Aoufous]] i/lub [[Ifezouane]])</small> | ||
| − | + | ??[[Niger]]<br> | |
| − | + | <small>(formacja [[Echkar]])</small> | |
| − | + | |czas = {{Występowanie|101|96}} | |
| − | + | 101-?96 [[Ma]]<br> | |
| − | + | późna [[kreda]] | |
| − | + | |systematyka = [[Dinosauria]] | |
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | <small>( | ||
| − | | | ||
| − | |||
| − | | | ||
| − | |||
| − | |||
| − | |||
| − | | | ||
| − | |||
| − | |||
[[Saurischia]] | [[Saurischia]] | ||
| Linia 54: | Linia 29: | ||
[[Spinosaurinae]] | [[Spinosaurinae]] | ||
| − | | | + | |grafika = Spinosaurus aegyptiacus 2015 by delirio88-d8e86yo.jpg |
| − | + | |podpis = Rekonstrukcja polującego w wodzie spinozaura.<br>Autor: Francesco Delrio | |
| − | + | [http://delirio88.deviantart.com/art/Spinosaurus-aegyptiacus-2015-507626304]. | |
| − | [http://delirio88.deviantart.com/art/Spinosaurus-aegyptiacus-2015-507626304]. | + | |PorównanieGrupa = Spinosauridae |
| − | | | + | |PorównanieDługość = 14 |
| − | + | |mapa = | |
| − | | | ||
| − | | | ||
<display_points type="terrain" zoom=3> | <display_points type="terrain" zoom=3> | ||
| − | 28.46024, 28.92426 | + | 28.46024, 28.92426~3 km na północ od Gebel el Dist (Stromer, 1914; 1915; Smith i in., 2006) - holotyp |
| − | 31°37N, 4°16W | + | 31°37N, 4°16W~Zrigat, 31°37N, 4°16W (Ibrahim i in., 2014) – FSAC-KK 11888 |
| − | 31.1228186,-4.8018245 | + | 31.1228186,-4.8018245~Okolice Taouz - w promieniu do ok. 200 km (Russell, 1996; Evers i in., 2015; Hendrickx i in., 2016) - okazy opisane przez Eversa i in. (2015) jako Spinosauridae [[indet]]. |
| − | 31.53407, -4.345093 | + | 31.53407, -4.345093~Okolica Jorf (Hendrickx i in., 2016) - MHNM.KK376 |
</display_points> | </display_points> | ||
| − | + | [[Plik:Geographical_location_and_stratigraphy_of_the_Kem_Kem_beds.PNG|350px|]] | |
| − | + | <small>Lokalizacja osadów Kem Kem (na czarno), w których odnaleziono okazy opisane przez Eversa i in. (2015). MHNM.KK378 został odnaleziony w pobliżu Jorf (zaznaczonego strzałką). Źródło: Hendrickx i in., 2016.</small> | |
| − | <small>Lokalizacja osadów Kem Kem (na czarno), w których odnaleziono okazy | + | }} |
| − | + | <small> | |
| − | opisane przez Eversa i in. (2015). MHNM.KK378 został odnaleziony w pobliżu Jorf (zaznaczonego | + | __TOC__ |
| − | + | </small> | |
| − | strzałką). Źródło: Hendrickx i in., 2016.</small> | ||
| − | |||
| − | |||
| − | |||
==Wstęp== | ==Wstęp== | ||
| − | ''Spinosaurus'' to [[rodzaj]] [[teropod]]a ([[spinozauryd]]a), który żył w "środkowej" kredzie na terenach obecnej północnej Afryki. Jedną z cech wyróżniających spinozaura były bardzo wysokie wyrostki kręgów grzbietowych | + | ''Spinosaurus'' to [[rodzaj]] [[teropod]]a ([[spinozauryd]]a), który żył w "środkowej" kredzie na terenach obecnej północnej Afryki. Jedną z cech wyróżniających spinozaura, od której wziął swoją nazwę, były bardzo wysokie wyrostki kolczyste kręgów grzbietowych. Opisanie nowego okazu w 2014 r., a także publikacja dodatkowych informacji w 2020 r. wskazuje, że był to teropod o niezwykłej budowie i biologii oraz pierwszy znany półwodny nieptasi dinozaur (Ibrahim i in., 2014). |
| − | ''Spinosaurus'' jest | + | ==Etymologia== |
| + | Nazwa ''Spinosaurus'' oznacza "kolczasty jaszczur" i jest zlepkiem łacińskiego słowa ''spina'', czyli "kolec", oraz greckiego ''sauros'', czyli "jaszczur" (Creisler, online 2003), i odnosi się do wysokich wyrostków kolczystych kręgów tego teropoda (Stromer, 1915). Epitet gatunkowy nawiązuje do Egiptu - kraju, w którym znaleziono jego pierwsze szczątki. | ||
| − | + | ''Spinosaurus'' dał nazwę nowej rodzinie [[Spinosauridae]] utworzonej przez Stromera w 1915 roku a później także nadrodzinie [[Spinosauroidea]] i podrodzinie [[Spinosaurinae]]. | |
| − | |||
==Filogeneza== | ==Filogeneza== | ||
| − | Zob. [[Spinosauridae# | + | Zob. [[Spinosauridae#Filogeneza i budowa]]. |
==Historia odkryć== | ==Historia odkryć== | ||
| − | ''Spinosaurus aegyptiacus'' to pierwszy [[rodzaj]] i [[gatunek]] egipskiego dinozaura, odkryty w osadach formacji [[Baharija]] [[1912]] roku i opisany przez niemieckiego paleontologa [[Ernst Stromer|Ernsta Stromera]] w [[1915]] r. Kolejne, dość fragmentaryczne, szczątki z tej formacji, składające się z kręgów i kości kończyn tylnych Stromer ([[1934]]) | + | [[Plik:Spinosaurus1.jpg|300px|thumb|right|Holotyp i rekonstrukcja szkieletu, pochodząca sprzed 2014 r. Autor: Scott Hartman [http://skeletaldrawing.blogspot.com/2012/01/revisiting-fisher-king.html].]] |
| + | [[Plik:Spinosaurus some cool dinosaurs by arvalis.png|300px|thumb|left|Rekonstrukcja sprzed 2014 r. Autor: RJ Palmer (arvalis) [http://arvalis.deviantart.com/art/Some-Cool-Dinosaurs-335980633].]] | ||
| + | |||
| + | ''Spinosaurus aegyptiacus'' to pierwszy [[rodzaj]] i [[gatunek]] egipskiego dinozaura, odkryty w osadach formacji [[Baharija]] [[1912]] roku i opisany przez niemieckiego paleontologa [[Ernst Stromer|Ernsta Stromera]] w [[1915]] r. Kolejne, dość fragmentaryczne, szczątki z osadów tej formacji, składające się z kręgów i kości kończyn tylnych, Stromer ([[1934]]) opisał jako ''„Spinosaurus B”''. Uznał, że mają wystarczająco wiele odmiennych cech, by utworzyć nowy [[gatunek]] w obrębie tego [[rodzaj]]u, lecz nie zdecydował się na to. Obecnie uznaje się, że przynajmniej część z tego materiału należała do innego [[spinozauryd]]a - ''[[Sigilmassasaurus]]'' (Evers i in., 2015). Oba okazy zostały zniszczone podczas nalotu bombowego na Monachium w czasie II wojny światowej, w nocy z 24 na 25 kwietnia 1944 r. | ||
| − | W [[1996]] roku [[Dale Russell]] opisał nowy [[gatunek]] - ''S. maroccanus'' - na podstawie kości odkrytych w marokańskich osadach [[Kem Kem]]. Za | + | W [[1996]] roku [[Dale Russell]] opisał nowy [[gatunek]] spinozaura - ''S. maroccanus'' - na podstawie kości odkrytych w marokańskich osadach [[Kem Kem]]. Za cechę odmienną uznał nieznacznie bardziej wydłużone kręgi szyjne. Jednak w dalszych pracach (Sereno i in., [[1998]]; Rauhut, 2003; Dal Sasso i in., [[2005]]) uznawano drugi gatunek za ''[[nomen dubium]]''. Różnice proporcji kręgów [[Michael Mortimer|Mortimer]] (online; cytowane przez Hendrickxa i in. [2016]) tłumaczył ich miejscem w kręgosłupie i [[synonim]]izował gatunek marokański z egipskim (ale zobacz dalej). |
| − | + | Ważny okaz - MSNM V4047 - przednia część czaszki - ujrzał światło dzienne w 2005 r. Skamieniałość pochodzi z Kem Kem w Maroko ([[Cristiano dal Sasso|dal Sasso]] i in., 2005). Jej identyfikacja jako ''Spinosaurus'' jest jednak wątpliwa (zob. dalej). | |
| − | Przełomowe znaczenie miała praca opublikowana w 2014 r. (Ibrahim i in., | + | Przełomowe znaczenie miała praca opublikowana w 2014 r. (Ibrahim i in.), w której pobieżnie opisano FSAC-KK 11888 - kolejny okaz z Maroka, obejmujący m.in. kości czaszki, kręgi oraz prawie wszystkie kości miednicy i kończyn tylnych. FSAC-KK 11888 rzucił nowe światło na budowę i tryb życia tego zwierzęcia, ukazując go jako półwodnego drapieżnika o skróconych kończynach tylnych. Publikacja wzbudziła wiele kontrowersji – zob. [[#Kontrowersje związane z FSAC-KK 11888]]. Ibrahim i in. (2014) uznali ''Spinosaurus maroccanus'' i ''[[Sigilmassasaurus]] brevicollis'' za [[synonim]]y ''Spinosaurus aegyptiacus''. Jednak rok później Evers i in. wykazali, że ''Sigilmassasaurus brevicollis'' to [[ważny]] [[gatunek]], odmienny od ''Spinosaurus aegyptiacus'', i dokonali jego [[synonim]]izacji ze ''Spinosaurus maroccanus''. Obecnie jedynym gatunkiem spinozaura jest więc [[gatunek typowy|typowy]] ''S. aegyptiacus''. |
| + | |||
| + | W 2020 r. Ibrahim i in. podtrzymali swoje zdanie na temat synonimizacji ''S. aegyptiacus'' z ''Sigilmassasaurus brevicollis'' i ''Spinosaurus maroccanus''. Ujawniono też nowe szczątki okazu FSAC-KK 11888, które po raz kolejny zmieniły nasze wyobrażenie o spinozaurze. Okazało się m.in. że ogon był znacznie wyższy niż dotąd sądzono. W tym samym roku Smyth i in. dokonali przeglądu niemal całego materiału kopalnego spinozaurydów z Kem Kem i nie znaleźli podstaw do wyrózniania dwóch taksonów, tym samym kwestionując odrębność sigilmassazaura. | ||
==Materiał kopalny== | ==Materiał kopalny== | ||
| − | [[Plik: | + | [[Plik:Spinosaurus aegyptiacus holotype skeletal.jpg|300px|thumb|right|Holotyp. Autor: Jaime A. Headden, zmodyfikowane przez IJReid [https://qilong.wordpress.com/2011/09/06/a-fistful-of-spinosaurus/], [https://commons.wikimedia.org/wiki/File:Spinosaurus_aegyptiacus_holotype_skeletal.jpg].]] |
===Holotyp=== | ===Holotyp=== | ||
| − | IPHG 1912 VIII 19 | + | [[Holotyp]] to IPHG 1912 VIII 19, opisany przez Stromera w 1915 r. i zniszczony podczas II wojny światowej. Okaz składał się z: fragmentu kości szczękowej, przedniej części żuchwy (kości zębowej, kości płatowej i ?kątowej), 15 izolowanych zębów, łuku neuralnego ?3 (Evers i in., 2015) [[kręg]]u szyjnego, 5(?4) (Evers i in., 2015) kręgu szyjnego, czterech (Stromer, 1915; Smith i in., 2006) trzonów kręgów grzbietowych i siedmiu wyrostków kolczystych, kilku żeber i [[gastralia|gastraliów]], trzech trzonów kręgów krzyżowych i jednego wyrostka kolczystego (Stromer, 1915). |
====Kontrowersje związane z holotypem==== | ====Kontrowersje związane z holotypem==== | ||
| − | + | Przypuszcza się, że kręg ogonowy wchodzący w skład holotypu pochodzi od innego [[takson]]u (Stromer, 1934; Rauhut, 2003, s. 35). W [[2003]] roku [[Oliver Rauhut]] zasugerował, że [[holotyp]] spinozaura był chimerą [[allozauroid]]a (kręgi grzbietowe) i teropoda podobnego do ''[[Baryonyx]]'' (żuchwa i zęby). Nie zostało to jednak przyjęte w późniejszych badaniach ([[Thomas Holtz|Holtz]] i in., [[2004]]; Dal Sasso i in., [[2005]]). W [[2015]] r. Evers i in. (w tym Rauhut) uznali, że skoro ''[[Ichthyovenator]]'' ma podobnie wydłużone wyrostki kręgów, to przychylają się do konsensusu panującego w tym względzie. Cau (online 2009) wskazał, że kręgi są najbardziej zbliżone do ''Baryonyx'' i ''[[Megaraptor]]'' (uważanego wtedy przez niego za [[spinozauryd]]a), co potwierdzać miało bliskość [[filogeneza|filogenetyczną]] kręgów i żuchwy. | |
| + | |||
| + | [[Plik:Img 6.jpg|300px|thumb|left|Rekonstrukcja holotypu. Elementy zrekonstruowane mają kolor szary. Skala = 1 m. Autor: Genya Masukawa (GetAwayTrike) [http://blogs.yahoo.co.jp/rboz_05/36950355.html].]] | ||
| + | [[Plik:Spinosaurus_FSAC-KK11888_GetAwayTrike.jpg|300px|thumb|left|Rekonstrukcja FSAC-KK 11888. Elementy zrekonstruowane mają kolor szary. Skala = 1 m. Autor: Genya Masukawa (GetAwayTrike) [http://blogs.yahoo.co.jp/rboz_05/36950355.html].]] | ||
===FSAC-KK 11888=== | ===FSAC-KK 11888=== | ||
| − | FSAC-KK 11888 jest bardziej zbliżony do [[holotyp]]u ''Spinosaurus'' niż do ''[[Sigilmassasaurus]]''. Dwa pierwsze mają charakterystyczne poszerzenia (“wybrzuszenia”) w dolnej części kręgów grzbietowych, co różni je od ''[[Sigilmassasaurus]]'' - ''„Spinosaurus B”'' (Stromer, 1934; Evers i in., 2015). | + | FSAC-KK 11888 jest bardziej zbliżony do [[holotyp]]u ''Spinosaurus'' niż do ''[[Sigilmassasaurus]]''. Dwa pierwsze okazy mają charakterystyczne poszerzenia (“wybrzuszenia”) w dolnej części kręgów grzbietowych, co różni je od okazu ''[[Sigilmassasaurus]]'' - ''„Spinosaurus B”'' (Stromer, 1934; Evers i in., 2015). Uzasadnia to przypisanie go do ''Spinosaurus''. |
FSAC-KK 11888 zawiera fragmenty łba, po kilka kręgów z każdej części kręgosłupa, dwa paliczki dłoni oraz niemal kompletne: obręcz miedniczną i kończyny tylne. | FSAC-KK 11888 zawiera fragmenty łba, po kilka kręgów z każdej części kręgosłupa, dwa paliczki dłoni oraz niemal kompletne: obręcz miedniczną i kończyny tylne. | ||
| − | Z publikacji Ibrahima i in. (2014) nie wynika jednoznacznie, które elementy, zwłaszcza kręgosłupa, są obecne. Jeśli nie podano inaczej, cytowanym źródłem jest Ibrahim i in. (2014); skróty odnoszą się do pozycji kręgów: C – szyjny (ang. cervical), D – grzbietowy (ang. dorsal), S – krzyżowy (ang. | + | [[Plik:Spinosaurus modified from Ibrahim et al, modified.PNG|300px|thumb|left|Rekonstrukcja szkieletu spinozaura Ibrahima i in. (2014). Elementy odnalezione, w tym niepewnie przypisane, mają kolor czerwony. Czarnymi kwadratami zaznaczono elementy holotypu, a czerwonymi - FSAC-KK 11888. Zob. „Uwaga do ilustracji” obok. Źródło: Hendrickx i in., 2016 (zmodyfikowane przez MZ) [http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0144695#pone-0144695-g011].]] |
| + | Z publikacji Ibrahima i in. (2014) nie wynika jednoznacznie, które elementy, zwłaszcza kręgosłupa, są obecne. Jeśli nie podano inaczej, cytowanym źródłem jest Ibrahim i in. (2014); skróty odnoszą się do pozycji kręgów: C – szyjny (ang. cervical), D – grzbietowy (ang. dorsal), S – krzyżowy (ang. sacral), Ca – ogonowyn (ang. caudal). Wydaje się, że w skład FSAC-KK 11888 wchodzą niżej wymienione kości. Część kości nosowej, kość przedczołowa, łuskowa, kwadratowo-jarzmowa (niezilustrowana?), większość obu kości kwadratowych, części kości zębowej, izolowane zęby, kość ?łzowa (suplement, s. 2-3). 2-?4 kręgi szyjne (podano wymiary dwóch trzonów, zidentyfikowanych jako C2 i C7 - suplement, s. 32 – tab. S2, lecz ryc. S3 sugeruje, że są trzy trzony - poza ww., też C3 i C9). Min. 5 trzonów kręgów grzbietowych (kręg widoczny na ryc. S2C to D1 wg ryc. S3, choć wg Eversa i in. [2015] kręgów szyjno-grzbietowych nie ma wśród odlewów FSAC-KK 11888 w University of Chicago; D4 i D5 są widoczne na ryc. S3; D5 – ryc. S2E i S3; D6, D7, D8 – wg suplementu s. 32 – tab. S2; wymiary podano tylko dla D6-D8), min. 1 kompletny wyrostek kolczysty (D8 – ryc. 2E) i kilka niekompletnych (Cau, online 2014A; ryc. S3 i ryc. w National Geographic [online 2014] sugerują, że elementy wyrostków zachowane w FSAC-KK 11888 to D4, D5, D6, D7 i D9; równie dobrze mogą one pochodzić z innej części kręgosłupa, np. krzyżowej). Trzony kręgów krzyżowych: S3, S4 i S5 (suplement, tab. 2). ?9 kręgów ogonowych (ryc. S3; nie podano wymiarów żadnego z nich). Żebra szyjne, grzbietowe i min. jedno brzuszne, min. 3 szewrony. Paliczek dłoni II-2 i bliższą część paliczka II-3. Kości miednicy i kończyn tylnych - brak kości pięty, paliczka II-2, kości śródstopia II i IV; nie wiadomo czy jest pazur I. | ||
| + | |||
| + | w 2018 r. zorganiowano kolejną ekspedycję, której owocem było odkrcie kolejnych kości okazu FSAC-KK 11888. Składa się na nie ponad 30 kręgów ogonowych, stanowiących ok. 80% całości ogona. odkryto też dodatkowe elementy czaszki i stopy (Ibrahim i in., 2020b). | ||
| + | |||
| + | <small>Uwaga do ilustracji. Z pokrywających się w holotypie i FSAC-KK 11888 elementów oznaczono te zilustrowane i niefragmentaryczne (np. z FSAC-KK 11888 znane są części kości zębowej). Identyfikacja kości łzowej wg Ibrahima i in. (2014) niepewna. Niezaznaczone są: zęby, żebra szyjne i grzbietowe oraz gastralia (wszystkie są obecne w obu okazach). Nie uwzględniono fragmentów wyrostków kolczystych (wg. ryc. S3 D4-7 i D8). Pominięto również zduplikowane elementy kończyn FSAC-KK 11888. Kości tylnej części czaszki nie zaznaczano osobno. Trzony kręgów szyjnych i grzbietowych i ich wyrostki kolczyste zaznaczono osobno: trzony pod nimi a wyrostki nad nimi. Trzony kręgów krzyżowych oznaczono jednak nad wyrostkami; pierwszy kręg zawiera w holotypie część wyrostka. Kręgi ogonowe, niezależnie od tego, czy zawierają trzony czy wyrostki, oznaczono nad nimi. | ||
| + | Identyfikacja elementów FSAC-KK 11888 wg Ibrahim i in., 2014 a holotypu wg Smith i in., 2006 i Stromer, 1915. Ryc. S3 z Ibrahim i in. (2014) jest mało przydatna ze względu na podobieństwo kolorów użytych do rozróżnienia obu okazów (o ile odróżnienie koloru żuchwy czy kości kończyn nie nastręcza trudności, to np. przy kręgach grzbietowych nie jest to proste; ponadto np. kość znajdująca się w tylnej części żuchwy – kątowa, choć jest wyraźnie oznaczona kolorem FSAC-KK 11888, należy do holotypu). | ||
| + | Rysunki z National Geographic, online 2014 nie odpowiadają w części ryc. S3 z Ibrahim i in. (2014) (np. tylny kręg szyjny FSAC-KK 11888 zakolorowany jest w całości, podczas gdy wyrostek kolczysty nie jest zakolorowany w ryc. S3; pominięto tylne kręgi szyjne i przednie grzbietowe; D6 – w National Geographic [online 2014] środkowa część wyrostka pochodzi od FSAC-KK 11888, tak jak w ryc. S3, lecz wg tej drugiej górna i dolna część wyrostka pochodzi od holotypu a w rzeczywistości holotyp ma cały wyrostek; zaznaczono tylko dwa kręgi ogonowe FSAC-KK 11888, podczas gdy w ryc. S3 widocznych jest 9, Ibrahim i in. [2014, s. 1613] podają, że FSAC-KK 11888 zawiera tylne kręgi ogonowe; 4-ty kręg ogonowy w holotypie jest kompletny a na ryc. S3 i w National Geographic [online 2014] wydaje się, że trzon jest z FSAC-KK 11888). | ||
| + | Kształt wyrostków kolczystych holotypu w znacznej mierze nie pokrywa się ze zdjęciem ze Smith i in., 2006 i rycinami ze Stromer, 1915. | ||
| + | Wyrostek kolczysty obecny w holotypie początkowo zidentyfikowano jako C2, lecz Evers i in. (2015) sugerują, że to C3. Ponieważ Ibrahim i in. (2014) podają wymiary trzonu C2, to przyjęto, że również wg nich kręg holotypu to C3 (również w National Geographic [online 2014] jest taka identyfikacja).</small> | ||
====Kontrowersje związane z FSAC-KK 11888==== | ====Kontrowersje związane z FSAC-KK 11888==== | ||
| − | [[Plik:Spinosaurus fishing by c compiler.jpg| | + | [[Plik:Spino potamus by chrismasna-d82kxwp.jpg|300px|thumb|right|Walczące ze sobą spinozaury. Autor: Chris Masna [http://chrismasna.deviantart.com/art/Spino-potamus-488065705].]] |
| − | + | [[Plik:Spinosaurus Marsh wader by julio lacerda.png|300px|thumb|right|Rekonstrukcja (w tym widoku budowa spinozaura raczej nie budzi kontrowersji). Autor: Julio Lacerda [http://julio-lacerda.deviantart.com/art/Marsh-Wader-400086688].]] | |
| + | [[Plik:Spinosaurus fishing by c compiler.jpg|300px|thumb|left|''Spinosaurus'' pożywiający się rybą (długość kończyn tylnych błędnie zawyżona - za Hartmanem, online 2014A). Autor: c-compiler [http://c-compiler.deviantart.com/art/Spinosaurus-Lunch-at-the-Beach-528724864].]] | ||
| − | + | Próba ustanowienia FSAC-KK 11888 [[neotyp]]em ''Spinosaurus aegyptiacus'' przez Ibrahima i in. (2014) została skrytykowana przez Eversa i in. (2015). Autorzy ci zwracają uwagę, że znaczna część tego okazu nie została odkryta przez Ibrahima i współpracowników, lecz odkupiona od prywatnego kolekcjonera. Część szkieletu została nabyta przez Uniwersytet w Casablance, a część przez Muzeum Historii Naturalnej w Mediolanie (Cau, online 2014A). Dopiero później zlokalizowano kolekcjonera, który doprowadził autorów opisu do miejsca znalezienia kości, dzięki czemu odkryto dalsze pozostałości (nie wiadomo jednak które). Brak więc danych o powiązaniu kości ze sobą. Evers i in. (2015) zwracają uwagę, że większość kręgowców z osadów [[Kem Kem]] jest znajdowanych w wielogatunkowych złożach. Zatem – jeśli nawet zostanie udowodnione, że cały materiał pochodzi z jednego stanowiska, to powinno się następnie wykazać, że są to szczątki jednego osobnika. Jako przesłanki za taką interpretacją autorzy wskazują zbliżoną wielkość i budowę kręgów oraz pasujące do siebie kości udowe i piszczele. Jednak kolekcjonerzy i handlarze skamieniałości z Maroka często kompletują szkielety, zawierające kości z różnych stanowisk, tak, aby do siebie pasowały. Tym samym nie można wykazać, że wszystkie kości wchodzące w skład FSAC-KK 11888 pochodzą od jednego osobnika (Evers i in., 2014). Jak jednak zauważają sami autorzy, występowanie podobnych nietypowych proporcji (znaczne skrócenie kończyn w porównaniu do kręgów) u dwóch okazów (FSAC-KK 11888 oraz u IPHG 1922 X45 - ''„Spinosaurus B”'', prawdopodobnie ''[[Sigilmassasaurus]]'') jest zastanawiające. Trudno uznać to za przypadek. Za pochodzeniem kości od jednego okazu świadczyć też mogą cechy młodociane obecne w wielu kościach (Ibrahim i in., 2014). Cau (online 2016A) sugeruje, że za hipotezą, że to jeden osobnik przemawia identyfikacja kości kwadratowej jako pochodzącej od młodego okazu (ang. subadult) przez Hendrickxa i in. (2016). Jednak kość FSAC-KK 11888 jest większa niż inne w tym samym i kolejnym stadium rozwoju, co świadczy przeciwko powiązaniu. | |
| − | Scott Hartman (online 2014A) oraz Mark Witton (online 2014B) wskazywali, że rekonstrukcja FSAC-KK 11888 przedstawia błędnie wyskalowaną miednicę i nogi spinozaura - wg nich są o 27% za małe. | + | W środowisku paleontologicznym nie ma pełnej zgody co do poprawności wniosków Ibrahima i in. (2014), opartych na budowie FSAC-KK 11888 oraz na będącej podstawą wielu ich hipotez rekonstrukcji, zawierającej szczątki tego oraz innych okazów. Scott Hartman (online 2014A) oraz Mark Witton (online 2014B) wskazywali, że rekonstrukcja FSAC-KK 11888 przedstawia błędnie wyskalowaną miednicę i nogi spinozaura - wg nich są o 27% za małe. Autorzy badania przekonująco wyjaśnili, że rozbieżności wynikają z innych punktów pomiaru w przypadku kręgów a w przypadku kości udowej także z jej niezupełnie pionowego ułożenia (Ibrahim, Maganuco, Keillor i Fabbri w Hartman, online 2014B; Maganuco w Witton, online 2014B). |
| − | |||
| − | Ibrahim i in. (2014) zaliczyli do ''Spinosaurus aegyptiacus'' szczątki znane jako ''„Spinosaurus B”'' (IPHG 1922 X45). Evers i in. (2015) wskazali jednak na różnice w budowie FSAC-KK 11888 | + | Ibrahim i in. (2014) zaliczyli do ''Spinosaurus aegyptiacus'' szczątki znane jako ''„Spinosaurus B”'' (IPHG 1922 X45). Za ich tożsamością mają świadczyć m.in. podobne proporcje długości kręgów do kończyn u IPHG 1922 X45 i FSAC-KK 11888 a także liczne podobieństwa, uwidocznione w ryc. S2 suplementu publikacji Ibrahima i in. (2014). Evers i in. (2015) wskazali jednak na różnice w budowie IPHG 1922 X45 i FSAC-KK 11888, dotyczące m.in. kończyn tylnych, i wykazali, że przynajmniej część tego materiału (kręgi) należy do ''[[Sigilmassasaurus]]'' (co było jednak później kwestionowane przez Ibrahima i in. [2020] oraz Smytha i in. [2020]). Jeśli więc IPHG 1922 X45 i FSAC-KK 11888 reprezentują ten sam takson a ten pierwszy różni się od holotypu spinozaura (konkretnie kręgów), to powstaje pytanie, czy ten ostatni faktycznie należy do ''Spinosaurus aegyptiacus'' (tak też wcześniej Mortimer, online 2014) (art. 75.3.5 [[International Code on Zoological Nomenclature|ICZN]]). Jak wyżej wskazano, FSAC-KK 11888 i holotyp spinozaura łączy budowa wyrostków kręgów, jak również mają inne cechy nieprzystające do materiału ''Sigilmassasaurus'' (Evers i in., 2015). Autorzy podkreślają znaczną odległość między miejscem znalezienia typowego okazu spinozaura i FSAC-KK 11888 (art. 75.3.6 ICZN), a także wyraźne różnice między [[Kem Kem]] (Maroko) i [[Baharija]] z (Egipt) (zob. też [[Carcharodontosaurus#Środowisko]]). Może się okazać, że FSAC-KK 11888 i inne szczątki spinozaurydów z Kem Kem, odmienne od ''[[Sigilmassasaurus]]'', reprezentują nowy, nienazwany jeszcze gatunek. Ponadto Evers i in., jak wcześniej Headden (online 2014), kwestionują konieczność wyznaczania [[neotyp]]u (art. 75.3 ''in principio'' ICZN), gdyż holotyp spinozaura, pomimo iż został zniszczony, to jest stosunkowo dobrze opisany i zilustrowany w literaturze a cechy diagnostyczne są widoczne. Ponadto są dostępne jego zdjęcia (Smith i in., 2006). |
Publikacja Ibrahima i in. (2014) budzi wiele innych zastrzeżeń. Jak wskazano wyżej, nie wiadomo, jaki materiał wchodzi w skład okazu (zob. wyżej - [[#FSAC-KK 11888]]). Autorzy nie podali, w jaki sposób mierzyli okazy ani jak wykonali rekonstrukcję, która posłużyła np. do wyznaczania środka masy zwierzęcia (np. Witton, online 2014A). | Publikacja Ibrahima i in. (2014) budzi wiele innych zastrzeżeń. Jak wskazano wyżej, nie wiadomo, jaki materiał wchodzi w skład okazu (zob. wyżej - [[#FSAC-KK 11888]]). Autorzy nie podali, w jaki sposób mierzyli okazy ani jak wykonali rekonstrukcję, która posłużyła np. do wyznaczania środka masy zwierzęcia (np. Witton, online 2014A). | ||
| + | [[Plik:Spinosaurus Evers2015 Fig-22-full.png|150px|thumb|right|Kręg szyjny (C?8) NHMUK PV R 36637. Źródło: Evers i in., 2015 [http://dx.doi.org/10.7717/peerj.1323].]][[Plik:Spinosaurus_Evers2015_Fig-17-full.png|200px|thumb|left|Kręg szyjny (C6) BSPG 2006 I 57. Wg Smytha i in. (2020) jest to kreg C5. Źródło: Evers i in., 2015 [http://dx.doi.org/10.7717/peerj.1323].]] | ||
| + | |||
| + | Ibrahim i in. (2020a) bronili swoich wniosków w kolejnej publikacji. Zwrócili uwagę, że FSAC-KK 11888 nie zawiera żadnych podwójnych kości, co powinno mieć miejsce, gdyby szczątki pochodziły od kilku zwierząt. Ponadto parzyste kości z obu stron ciała mają w zasadzie identyczne rozmiary i budowę, a część kości dłoni zachowała się w naturalnym ułożeniu. Ibrahim i współpracownicy uznali również, że stan zachowania kości wskazuje, że przynajmniej część miednicy i kończyn tylnych także była artykułowana w momencie ich znalezienia. Bronili oni także zasadności wyznaczenia okazu FSAC-KK 11888 na neotyp. | ||
| + | |||
| + | Publikacja Ibrahima i in. (2020b) ujawniła dodatkowy materiał okazu FSAC-KK 11888, co w opinii autorów wzmocniło ich konkluzje na temat tego, że szczątki te pochodziły od jednego osobnika. Dzięki odkryciu dodatkowych kręgów ogonowych, możliwe było porównanie z materiałem "''Spinosaurus'' B". Wykazało ono bardzo duże podobieństwo obu okazów pod względem budowy kręgów ogonowych. | ||
| + | |||
| + | Smyth i in. (2020) również postulowali zasadność wyznaczenia neotypu i uznali FSAC-KK 11888 za pewny materiał ''Spinosaurus aegyptiacus''. Kwestionowali m.in. podnoszony przez Eversa i współpracowników argument odległości geograficznej, gdyż ci sami autorzy sugerowali, że część materiału "''Spinosaurus B''" (Egipt) należy do sigilmassazaura, którego holotyp pochodzi z Maroka. | ||
===Materiał przypisany=== | ===Materiał przypisany=== | ||
| − | + | Badania Eversa i in. (2015), Hendrickxa i in. (2016) oraz Ardena i in. (2018) wskazują, że w [[cenoman]]ie na obszarze dzisiejszej Afryki Północnej – zarówno w Maroku, jak i w Egipcie, żyły dwa gatunki [[spinozauryd]]ów – prawdopodobnie ''Spinosaurus aegyptiacus'' i ''[[Sigilmassasaurus]] brevicollis''. Oba są znane fragmentarycznie, więc trudno oznaczyć większość znalezisk. Chiarenza i [[Andrea Cau|Cau]] (2016) uznali, że jako pewny materiał ''Spinosaurus aegyptiacus'' powinno się traktować wyłącznie [[holotyp]], a szczątki spinozaurydów z cenomanu Afryki Północnej, których nie można niewątpliwie przypisać do któregoś z dwóch taksonów, powinny być przypisywane tylko do [[Spinosaurinae]] do czasu gruntownej rewizji ich taksonomii. | |
| + | [[Plik:Spinosaurus_aegyptiacus.png|300px|thumb|left|Rekonstrukcja płynącego spinozaura. Autor: Gustavo Monroy-Becerril. [https://commons.wikimedia.org/wiki/File:Spinosaurus_aegyptiacus.png].]] | ||
| − | + | Prawdopodobnie wszystkie poniższe szczątki pochodzą z osadów [[Kem Kem]] z Maroka: | |
| − | * Cztery kręgi opisane przez Eversa i in. (2015): NHMUK PV R 16429 – przedni kręg szyjny, BSPG 2006 I 57 i BSPG 2013 I 97 – dwa środkowe kręgi szyjne oraz NHMUK PV R 36637 – tylny środkowy kręg szyjny. Autorzy wspomnieli też o okazie ROM 65537 – środkowym kręgu szyjnym. BSPG 2006 I 57 ma cechy zbliżające go do FSAC-KK 11888 (Evers i in., | + | * Cztery kręgi opisane przez Eversa i in. (2015): NHMUK PV R 16429 – przedni kręg szyjny, BSPG 2006 I 57 i BSPG 2013 I 97 – dwa środkowe kręgi szyjne oraz NHMUK PV R 36637 – tylny środkowy kręg szyjny. Autorzy wspomnieli też o okazie ROM 65537 – środkowym kręgu szyjnym. Evers i in. (2015) określali je jako „kręgi drugiego spinozauryda z Kem Kem” a ponieważ różnią się od ''[[Sigilmassasaurus]]'', to prawdopodobnie należą do ''Spinosaurus''. BSPG 2006 I 57 ma cechy zbliżające go do FSAC-KK 11888 (Evers i in., 2015). |
* Dwa środkowe kręgi szyjne, opisane początkowo jako [[paratyp]]y ''S. maroccanus'' (CMN 41768, CMN 50790). Tak samo jak powyższe, Evers i in. (2015) uznali je za bliżej nieokreślonego spinozauryda. Mortimer zaliczył je do ''Spinosaurus'' (tj. jako „aegyptiacus morphotype”). Jako że różnią się one od ''[[Sigilmassasaurus]]'' (Russell, 1996, ryc. 4d i 7), powinno się je przypisać do ''Spinosaurus''. | * Dwa środkowe kręgi szyjne, opisane początkowo jako [[paratyp]]y ''S. maroccanus'' (CMN 41768, CMN 50790). Tak samo jak powyższe, Evers i in. (2015) uznali je za bliżej nieokreślonego spinozauryda. Mortimer zaliczył je do ''Spinosaurus'' (tj. jako „aegyptiacus morphotype”). Jako że różnią się one od ''[[Sigilmassasaurus]]'' (Russell, 1996, ryc. 4d i 7), powinno się je przypisać do ''Spinosaurus''. | ||
| − | * Pięć kości kwadratowych, opisanych przez Hendrickxa i in. (2016), pochodzących od osobników w różnych fazach rozwoju osobniczego (MHNM.KK374, MHNM.KK375, MHNM.KK377, MHNM.KK378 oraz MSNM V6896). | + | * Pięć kości kwadratowych, opisanych przez Hendrickxa i in. (2016), pochodzących od osobników w różnych fazach rozwoju osobniczego (MHNM.KK374, MHNM.KK375, MHNM.KK377, MHNM.KK378 oraz MSNM V6896). Hendrickx i in. (2016) zaliczyli je do spinozaura ze względu na większe podobieństwo do FSAC-KK 11888 niż do drugiego [[morfotyp]]u kości kwadratowych z Kem Kem. |
| + | |||
| + | * Pazur stopy bardzo młodego osobnika (MSNM V6894), mierzącego ok. 1,8 m, który bardzo mocno przypomina pazury FSAC-KK 11888 (Maganuco i Dal Sasso, 2018). | ||
| + | |||
| + | * Dwie kości czołowe, w tym prawa niekompletna (FSAC-KK-3209); dwie kości czołowe i ciemieniowe (FSAC-KK-3210) wstępnie przypisane do ''Spinosaurus'' przez Ardena i in. (2018). Reprezentują one mniejszy z dwóch morfotypów wyróżnionych przez autorów. Wg Ibrahima i in. (2020a) nie należą one do spinozaurydów, lecz najprawdopodobniej do krokodylomorfów. | ||
| + | |||
| + | *Kręg szyjny C5 lub C6 (FSAC-KK-7280) oraz przedni kręg grzbietowy (FSAC-KK-18118) przypisane przez Lakin i Longricha (2018). Oba kręgi nie wykazują cech diagnostycznych ''[[Sigilmassasaurus]]'', mają za to cechy właściwe dla holotypu spinozaura. | ||
| + | |||
| + | *Sześć kręgów oznaczonych numerami FSAC-KK-5017 do FSAC-KK-5022 (pierwsze pięć to przednie i środkowe kręgi szyjne, ostatni - przedni grzbietowy, prawdopodobnie D2), przypisanych przez Smytha i in. (2020). | ||
===Materiał wątpliwy=== | ===Materiał wątpliwy=== | ||
| − | [[Plik:Journal.pone.0144695.g011.PNG| | + | [[Plik:Journal.pone.0144695.g011.PNG|150px|thumb|right|Pyski nieoznaczonych [[spinozauryn]]ów z cenomanu Afryki Północnej (''Spinosaurus'' lub ''[[Sigilmassasaurus]]''). Widoki A-B, E i G: MSNM V4047. Widoki C-D, F i H: MNHM SAM 124. Źródło: Hendrickx i in., 2016 [http://dx.doi.org/10.1371/journal.pone.0144695.g011].]] |
| − | W przeszłości wiele okazów spinozaurydów zaliczano do ''Spinosaurus'', lecz mogą należeć równie dobrze do ''[[Sigilmassasaurus]]''. | + | W przeszłości wiele okazów spinozaurydów zaliczano do ''Spinosaurus'', lecz obecnie wiadomo, że mogą należeć równie dobrze do ''[[Sigilmassasaurus]]''. Są to głównie fragmenty czaszki oraz liczne zęby. Najważniejszym z nich jest MSNM V4047 (kości przedszczękowe i niekompletne kości szczękowe i nosowe), opisany przez włoskiego paleontologa [[Cristiano Dal Sasso]] z Muzeum Historii Naturalnej w Mediolanie i innych naukowców w 2005 roku. MSNM V4047 jest największym jak dotąd odkrytym osobnikiem spinozauryda (Dal Sasso i in., 2005; Hednrickx i in., 2016) (zob. jednak [[Sigilmassasaurus#Rozmiary]]). Sama czaszka mierzyła według Dal Sasso i in. 1,75 metra, a całe zwierzę miało być może około 16-18 metrów długości. W rzeczywistości czaszka mogła być znacznie krótsza - zob. artykuł o [[Spinosauridae#Wymiary]]. Ponadto MSNM V4047 ma cofnięte do tyłu nozdrza a prześwietlenie wykazało, że prawdopodobnie pysk mieścił receptory ciśnienia wody (Ibrahim i in., 2014), zob. [[Spinosauridae#Paleobiologia]]. Arden i in. (2018) uznali za bardzo prawdopodobne, że okaz ten należał do ''[[Sigilmassasaurus]]'', ich zdaniem większego z dwóch spinozaurydów z cenomanu Afryki Północnej. Wg Lacerdy i in. (2021) MSNM V4047 na chwilę obecną nie może być definitywnie przypisywany do ''Spinosaurus''. |
| + | |||
| + | Do ''Spinosaurus'' może też należeć kość kwadratowa (FSAC-KK-18120) opisana przez Lakin i Longricha (2018). Reprezentuje ona morfotyp, który Hendrickx i in. (2016) zaliczyli do omawianego tu rodzaju. Lakin i Longricha nie zdecydowali się jednak na przypisanie jej do konkretnego spinozauryda z powodu braku materiału czaszkowego w holotypie spinozaura oraz niejasności związanych z FSAC-KK 11888. Podobnie postąpili z kręgiem szyjnym (FSAC-KK-18119) oraz przednim kręgiem piersiowym (FSAC-KK-18121), które wykazują cechy zarówno ''Spinosaurus'' jak i ''[[Sigilmassasaurus]]''. Ponadto, jako Spinosauridae indet. autorzy ci opisali prawą kość przedszczękową (FSAC-KK-7281), która przypomina okaz MSNM V4047, mogący należeć do sigilmassazaura. Podobnego zdania są Lacerda i in. (2021). | ||
| + | |||
| + | Spinozaurowi przypisywano też szczątki z cenomańskiej formacji [[Echkar]] w Nigrze. Należy to nich kręg grzbietowy, początkowo przypisany do ''[[Carcharodontosaurus]] iguidensis'', fragmentaryczny pysk (MNBH EGA1) oraz pojedyncze zęby. Sereno i in. (2022) zaznaczają, że szczątki te przypominają materiał ''S. aegyptiacus'', lecz mogą należeć do innego gatunku. Równie prawdopodobne wydaje się, że może to być odrębny rodzaj spinozauryda. | ||
| + | |||
| + | Najnowsze publikacje (Ibrahim i in., 2020a; Ibrahim i in., 2020b; Smyth i in., 2020) wskazują na synonimikę ''[[Sigilmassasaurus]]'' i ''Spinosaurus'', a różnice między nimi uznaje się w nich za wynik zmienności osobniczej, możliwego dymorfizmu płciowego, ontogenezy, pośmiertnych uszkodzeń szczątków, różnego stanu ich zachowania oraz porównywania ze sobą różnych kości, zwłaszcza kręgów, które mogły cechować się zmienną morfologią w zależności od ich położenia w kręgosłupie, nawet u pojedynczego osobnika. Tym samym cały materiał przypisywany do sigilmassazaura należałby do ''Spinosaurus''. Na obecną chwilę wydaje się, że kwestia ta wymaga jeszcze dalszych badań. | ||
==Budowa i paleobiologia== | ==Budowa i paleobiologia== | ||
===Plan ciała, budowa i paleobiologia ogólnie=== | ===Plan ciała, budowa i paleobiologia ogólnie=== | ||
| − | Czaszka była prawdopodobnie wydłużona | + | [[Plik:Spinosaurus aegyptiacus by camusaltamirano.jpg|300px|thumb|left|Rekonstrukcja. Autor: Camus Altamirano [http://camusaltamirano.deviantart.com/art/Spinosaurus-aegyptiacus-368230400].]] |
| + | Czaszka ''Spinosaurus'' była prawdopodobnie wydłużona, jak u innych [[spinozauryd]]ów (zob. niżej - [[#Czaszka]]). W przeciwieństwie do nich, długi był też tors, na którym znajdował się wysoki grzebień (zob. niżej - [[#Żagiel]]). O ogonie niewiele można stwierdzić – jak u innych [[teropod]]ów, był zapewne długi i silnie umięśniony. Opierając się na krewniakach, uznać należy, że kończyny przednie były zapewne dobrze rozwinięte i pomocne w łapaniu ryb. Ibrahim i in. (2014) wspomnieli, że dłonie były proporcjonalnie większe niż u wcześniejszych spinozaurydów, nie podali jednak dokładniejszych danych. Kończyny tylne były zredukowane i słabo przystosowane do dźwigania ciężaru ciała (zob. niżej - [[#Kończyny tylne]]). | ||
| − | Ibrahim i współpracownicy (2014) dowiedli, że co najmniej kości udowe i wyrostki żagla FSAC-KK 11888 były zbudowane głównie z gęstej tkanki kostnej, ze zmniejszoną jamą szpikową. Gęstość kości | + | Ibrahim i współpracownicy (2014) dowiedli, że co najmniej kości udowe i wyrostki żagla FSAC-KK 11888 były zbudowane głównie z gęstej tkanki kostnej, ze zmniejszoną jamą szpikową. Gęstość kości była 30-40% większa niż u innych teropodów i podobna jak u pingwinów królewskich. Interpretuje się to jako przystosowanie do półwodnego trybu życia (Ibrahim i in., 2014). |
| − | ''Spinosaurus'' był niezwykłym zwierzęciem, diametralnie różnym nie tylko od innych teropodów, ale także pozostałych [[spinozauryd]]ów. Dotyczy to zwłaszcza nietypowych proporcji kończyn tylnych i jego domniemanej czworonożności, czym różnił się choćby od ''[[Suchomimus]]''. Mimo pewnych zastrzeżeń (zob. [[#Kontrowersje związane z FSAC-KK 11888]]), trudno zaprzeczyć, że ''Spinosaurus'' był dobrze przystosowany do wodnego trybu życia i rybożerności | + | ''Spinosaurus'' był niezwykłym zwierzęciem, diametralnie różnym nie tylko od innych teropodów, ale także pozostałych [[spinozauryd]]ów. Dotyczy to zwłaszcza nietypowych proporcji kończyn tylnych i jego domniemanej czworonożności, czym różnił się choćby od ''[[Suchomimus]]''. Mimo pewnych zastrzeżeń (zob. [[#Kontrowersje związane z FSAC-KK 11888]]), trudno zaprzeczyć, że ''Spinosaurus'' był dobrze przystosowany do wodnego trybu życia i rybożerności. |
| − | + | W tyn kontekście zaskakujące są wyniki badań składu chemicznego zębów. Wynika z nich, że przedstawiciele Spinosauridae spędzali w wodzie znaczną ilość czasu, podobnie jak krokodyle czy hipopotamy. Jednak [[spinozauryd]]y z [[cenoman]]u Afryki Północnej – w tym na pewno ''Spinosaurus'' – okazały się bardziej lądowe niż ich krewniacy (Amiot i in., 2010). Zob. też [[Spinosauridae#Paleobiologia]]. | |
===Diagnoza=== | ===Diagnoza=== | ||
| − | Ibrahim i in. (2014) podali 10 pozaczaszkowych cech diagnostycznych spinozaura (literą „H” i „F” oznaczono cechy obecne odpowiednio w holotypie i | + | [[Plik:Spinosaurus holotype.jpg|300px|thumb|left|Elementy holotypu (1 – kręg ogonowy, 2, 4 – gastralia, 3 – lewa kość kątowa, 5, 7-11 – zęby, 6, 12 – żuchwa, 13-15 – żebra, 16 – kręgi krzyżowe, 17-19 - kręgi grzbietowe). Widoczny m.in. charakterystyczny kształt żuchwy. Źródło: Stromer, 1915 [https://www.flickr.com/photos/internetarchivebookimages/16531204169].]] |
| + | [[Plik:Spinosaurus_Stromer1915.jpg|200px|thumb|right|Elementy holotypu (1 i 2 - kręgi szyjne, 3-6 – kręgi grzbietowe). Widoczny m.in. charakterystyczny kształt kręgów grzbietowych. Źródło: Stromer, 1915 [https://www.flickr.com/photos/internetarchivebookimages/16531204169].]] | ||
| + | |||
| + | Ibrahim i in. (2014) podali 10 pozaczaszkowych cech diagnostycznych spinozaura (literą „H” i „F” oznaczono cechy obecne odpowiednio w holotypie i u FSAC-KK 1188, za odpowiednio Stromerem [1915] i Ibrahimem i in. [2014]). | ||
| + | |||
| + | (1) Silnie zwężone i wydłużone trzony kręgów grzbietowych o kształcie klepsydry (H, F [Cau, online 2014A]) – powodowało to znaczne wydłużenie grzbietu zwierzęcia. | ||
Kolejne cechy odnoszą się do tworzących „żagiel” wyrostków kolczystych kręgów grzbietowych, które: (2) były do 10 razy wyższe niż wysokość trzonów (H, F); (3) miały największą przednio-tylną szerokość poniżej swych wierzchołków (ang. „below spine apex”) (sic!) – być może chodzi o unikalne rozszerzenia (wypukłości) widoczne w dolnej części wyrostków (H, F); (4) były zbudowane z gęstej tkanki kostnej z wąskim pasem tkanki gąbczastej (H, F); (5) były pokryte pionowymi prążkami – w diagnozie (suplement, s. 9) podano, że znajdują się one na bliższej (dolnej) jedną trzeciej wyrostków, lecz w publikacji (s. 1616 i ryc. 4D) wskazano, że są one na dystalnej (górnej) powierzchni; w niektórych kręgach holotypu widoczne jest to na całej wysokości (H, F). | Kolejne cechy odnoszą się do tworzących „żagiel” wyrostków kolczystych kręgów grzbietowych, które: (2) były do 10 razy wyższe niż wysokość trzonów (H, F); (3) miały największą przednio-tylną szerokość poniżej swych wierzchołków (ang. „below spine apex”) (sic!) – być może chodzi o unikalne rozszerzenia (wypukłości) widoczne w dolnej części wyrostków (H, F); (4) były zbudowane z gęstej tkanki kostnej z wąskim pasem tkanki gąbczastej (H, F); (5) były pokryte pionowymi prążkami – w diagnozie (suplement, s. 9) podano, że znajdują się one na bliższej (dolnej) jedną trzeciej wyrostków, lecz w publikacji (s. 1616 i ryc. 4D) wskazano, że są one na dystalnej (górnej) powierzchni; w niektórych kręgach holotypu widoczne jest to na całej wysokości (H, F). | ||
| − | Diagnostyczna jest także budowa kończyn tylnych | + | Diagnostyczna jest także budowa kończyn tylnych: |
| − | (6) kości długie - tj. co najmniej udowa - nie miały otwartej jamy szpikowej (F); (7) długość kości biodrowej większa niż długość kości udowej (F); (8) kość udowa silnie wygięta ku przodowi (ang. „bowed anteriorly”), z rozrośniętym czwartym krętarzem, rozciągającym się wzdłuż 25% trzonu tej kości (F); (9) | + | (6) kości długie - tj. co najmniej udowa - nie miały otwartej jamy szpikowej (F); (7) długość kości biodrowej była większa niż długość kości udowej (F); (8) kość udowa była silnie wygięta ku przodowi (ang. „bowed anteriorly”), z rozrośniętym czwartym krętarzem, rozciągającym się wzdłuż 25% trzonu tej kości (F); (9) pierwszy palec stopy był długi; paliczek I-1 był najdłuższy z (niepazurowych) paliczków stopy (F); (10) pazury stóp były szersze niż ich głębokość (wysokość), o długości prawie cztery razy większej od bliższej głębokości; pazury miały płaską dolną powierzchnię (F). |
| − | Cechy te związane były prawdopodobnie w znacznej mierze z lokomocją (2?, 3?, 7 | + | Cechy te związane były prawdopodobnie w znacznej mierze z lokomocją (2?, 3?, 7, 8, 9, 10) i z wodnym trybem życia (1?, 3?, 4, 6, 7, 8, 10). |
Ibrahim i in. (2014) wymienili także dwie cechy pyska, jednak nie odnoszą się one do niewątpliwego materiału ''Spinosaurus''. | Ibrahim i in. (2014) wymienili także dwie cechy pyska, jednak nie odnoszą się one do niewątpliwego materiału ''Spinosaurus''. | ||
| − | Kilka „potencjalnie [[autapomorfia|autapomorficznych cech]]” obecnych w holotypie wymienili także Evers i in. (2015, s. 74). | + | |
| − | + | Kilka „potencjalnie [[autapomorfia|autapomorficznych cech]]” obecnych w holotypie wymienili także Evers i in. (2015, s. 74). | |
| − | ''Spinosaurus'' z pewnością jest odróżnialny od sigilmassazaura – różni się od niego prawdopodobnie budową kręgów i kończyn (Evers i in., 2015) oraz kości kwadratowej (Hendrickx i in., 2016) (zob. też [[Sigilmassasaurus#Budowa i paleobiologia]]). | + | ''Spinosaurus'' z pewnością jest odróżnialny od sigilmassazaura – różni się od niego prawdopodobnie budową kręgów i kończyn (Evers i in., 2015) oraz kości kwadratowej (Hendrickx i in., 2016) (zob. też [[Sigilmassasaurus#Budowa i paleobiologia]]). Odmienna była też budowa kości czołowych – te przypisane spinozaurowi były proporcjonalnie dłuższe i węższe (Arden i in. 2018). Ibrahim i in. (2020a) uznali jednak, że kości te nie należały w ogóle do spinozaurydów. Z kolei Smyth i in. (2020) uznali różnice w budowie kręgów i kości kwadratowej za nieistotne taksonomicznie. |
===Czaszka=== | ===Czaszka=== | ||
| − | [[Plik:Spinosaurus | + | [[Plik:Spinosaurus and onchopristis by alexanderlovegrove-d91tfgi.jpg|200px|thumb|right|Spinozaur polujący na rybę z rodzaju Onchopristis Autor: Alexanderlovegrove [http://whhl.deviantart.com/art/Spinosaurus-and-Onchopristis-547248258].]] |
| − | Czaszka spinozaura jest znana jedynie z kilku elementów. W zasadzie jedyny pewny a niefragmentaryczny materiał to część żuchwy. Prawdopodobnie miał on długą i niską czaszkę, z przesuniętymi do tyłu nozdrzami i stożkowatymi zębami. Czaszka niezidentyfikowanego spinozauryda MSNM V4047 ma na pysku liczne otwory i kanały – prawdopodobnie mieszczące receptory ciśnienia wody, zob. [[Spinosauridae#Paleobiologia]]. | + | |
| + | Czaszka spinozaura jest znana jedynie z kilku elementów. W zasadzie jedyny pewny a niefragmentaryczny materiał to część żuchwy. Prawdopodobnie miał on długą i niską czaszkę, z przesuniętymi do tyłu nozdrzami i stożkowatymi zębami. Czaszka niezidentyfikowanego spinozauryda MSNM V4047 ma na pysku liczne otwory i kanały – prawdopodobnie mieszczące receptory ciśnienia wody, zob. [[Spinosauridae#Paleobiologia]]. Wg Ardena i in. (2018) nietypowa budowa kości czołowych wskazuje na wysokie umiejscowienie oczodołów, ponad resztę czaszki (z czym jednak nie zgodzili się Hone i Holtz [2021]) oraz zredukowane opuszki węchowe. Te ostatnie prawdopodobnie były mało przydatne dla zwierzęcia polującego głównie w wodzie. | ||
| + | |||
| + | Niekompletna żuchwa została opisana przez Stromera (1915). Grzbietowa (górna) krawędź tworzy łuk, wypukły do szóstego zębodołu, dalej ku tyłowi wklęsły do dwunastego zębodołu, który następnie znów staje się wypukły i lekko wznosi się do góry. Brzuszna (dolna) krawędź żuchwy ma kształt zwróconego przeciwnie łuku, który najwyższą wysokość osiąga w okolicach siedemnastego zębodołu. Końcówka żuchwy jest kanciasta i stosunkowo wysoka, nadając jej specyficzny kształt. Pomiędzy trzecim a czwartym zębodołem żuchwa ma 13,5 cm wysokości, w okolicach siódmego – zaledwie 9 cm, na wysokości piętnastego – 15,5 cm, a 10 cm dalej ku tyłowi – aż 19 cm. Jej budowa wskazuje, że ''Spinosaurus'' miał długi, wąski pysk. Wg Stromera, w tylnej części kości żuchwy wydają się „cienkie i słabe”. Co ciekawe, obie połówki dolnej szczęki różnią się ilością zębów – prawa ma 15 zębodołów, a lewa – 16. Zęby mocno różniły się wielkością w zależności od położenia w żuchwie. Zęby od drugiego do czwartego, a więc leżące w podniesionej przedniej części żuchwy były wyraźnie większe od pozostałych, po nich następuje seria mniejszych zębów aż do dziesiątego, kolejne cztery znowu stają się większe, a ostatni jest ponownie mniejszy i silniej bocznie spłaszczony. Zęby spinozaura były tylko nieznacznie zakrzywione, a w przekroju poprzecznym – prawie okrągłe. Ich korzenie były niezwykle długie i w górnej części grubsze od koron. | ||
| − | + | W 2023 r. Smith i Martill opisali niezwykły patologicznie zmieniony ząb z osadów Kem Kem, mogący należeć do spinozaura. Wykazuje on m.in. zwiększoną krzywiznę korony i obecność trzech głębokich rowków wzdluż całej korony. | |
===Kończyny tylne=== | ===Kończyny tylne=== | ||
| − | Budowa kończyn tylnych wg Ibrahima i in. (2014) wskazuje na przystosowanie do półwodnego trybu życia. | + | [[Plik:Spinosaurus_hip.jpg|300px|thumb|left|Rekonstrukcja szkieletu wg Ibrahima i in. (2014). Fot.: Mike Bowler [https://www.flickr.com/photos/mbowler/15116479538/].]] |
| − | + | Budowa kończyn tylnych wg Ibrahima i in. (2014) wskazuje na przystosowanie do półwodnego trybu życia. Były one skrócone a miednica była zredukowana. Jak jednak sugeruje Cau (online 2016C), nie tyle kończyny były pomniejszone, co wydłużony był grzbiet. | |
| − | |||
| − | Wydłużenie pierwszego palca stopy (zredukowanego u innych teropodów) i niskie, spłaszczone pazury (podobne do pazurów ptactwa brzegowego) mogły być przystosowaniem do poruszania się po grząskim gruncie i wiosłowania w wodzie. Być może między palcami znajdowała się błona pławna. | + | W przeciwieństwie do innych dużych teropodów, kość udowa była krótsza od piszczelowej. Stosunkowo długie było także śródstopie. Pod względem proporcji długości kończyn ''Spinosaurus'' wydaje się najlepiej przystosowany do szybkiej lokomocji wśród bazalnych [[tetanur]]ów. Współczynnik obliczony metodą Personsa i Curriego (2016) na podstawie wymiarów podanych przez Ibrahima i in. (2014) to +8%. Wśród nie[[celurozaur]]owych [[teropod]]ów jedynie niektóre [[ceratozaury]] go przewyższają (zob. [[Ceratosauria#Lokomocja]]). U teropodów proporcje te zmieniają się znacznie w trakcie wzrostu – u młodych kończyny tylne, zwłaszcza dolne części, są proporcjonalnie dłuższe (np. Persons i Currie, 2016); u spinozaura mogło być podobnie. Takie proporcje u ''Spinosaurus''z pewnością nie znamionują jednak szybkiego ruchu, biorąc pod uwagę jak niewielkie są tylne kończyny w porównaniu do szkieletu osiowego (Ibrahim i in., 2014; jak wskazano [[#Kontrowersje związane z FSAC-KK 118888|wyżej]] nie ma jednak pewności co do pochodzenia szczątków od jednego osobnika). Kość udowa miała masywne miejsca przyczepu mięśni odpowiedzialnych za zginanie kończyny do tyłu. Dalsze kłykcie kości udowej są wąskie a grzebień goleniowy piszczeli średnio rozwinięty. Taka budowa przypomina stan obserwowany u wczesnych waleni i współczesnych półwodnych ssaków, używających łap do wiosłowania. Wydłużenie pierwszego palca stopy (zredukowanego u innych teropodów) i niskie, spłaszczone pazury (podobne do pazurów ptactwa brzegowego) mogły być przystosowaniem do poruszania się po grząskim gruncie i wiosłowania w wodzie. Być może między palcami znajdowała się błona pławna. Co ciekawe, budowa i proporcje pazurów pozostawały prawdopodobnie takie same przez całe życie, ułatwiając pokonywanie miękkich podłoży lub wiosłowanie. Wskazuje to, że tryb życia młodych spinozaurów nie odbiegał zbytnio od dorosłych (Maganuco i Dal Sasso, 2018). Potwierdzają to Lakin i Longrich (2018) którzy sugerują jednak, że bardzo młode osobniki (poniżej 2 m długości) zajmowały nieco inne nisze ekologiczne i występowały w innym środowisku. |
Proporcje ciała spinozaura – wydłużony grzbiet i krótkie kończyny – są częste u zwierząt wodnych (Nash, online 2014B). Zbliżoną budowę miał prawdopodobnie bliski krewny spinozaura - ''[[Sigilmassasaurus]]'' (Evers i in., 2015). | Proporcje ciała spinozaura – wydłużony grzbiet i krótkie kończyny – są częste u zwierząt wodnych (Nash, online 2014B). Zbliżoną budowę miał prawdopodobnie bliski krewny spinozaura - ''[[Sigilmassasaurus]]'' (Evers i in., 2015). | ||
| − | |||
===Żagiel=== | ===Żagiel=== | ||
====Budowa żagla==== | ====Budowa żagla==== | ||
| − | + | [[Plik:Spinosaurus swimming.jpg|300px|thumb|left|Szkielet ''Spinosaurus'' zrekonstruowany w pozycji płynącej. Autor zdjęcia: Mike Bowler [https://www.flickr.com/photos/mbowler/15116509577].]] | |
| + | |||
| + | Tzw. "żagiel" spinozaura był niezwykły, choć inne dinozaury, takie jak [[ornitopod]] ''[[Ouranosaurus]]'' czy [[hadrozauryd]] ''[[Barsboldia]]'', wytworzyły podobne struktury kostne. Podobne twory miały także permskie synapsydy, takie jak ''Dimetrodon'' i ''Edaphosaurus''. Żagiel jest cechą wytworzoną niezależnie u każdego spośród wymienionych gadów - czyli analogiczną. Wyrostki spinozaura były też najdłuższe wśród dinozaurów – miały do ok. 165 cm wysokości i były do 8 (Stomer, 1915) lub 10 (Ibrahim i in., 2014) razy wyższe od trzonów. | ||
| − | Kształt ozdoby jest niepewny. Tradycyjnie uznawano, że żagiel rozciągał się półkoliście nad grzbietem (Smith i in., 2006). Proponowano również, że najdłuższy wyrostek kolczysty znajdował się na ogonie (Cau, online 2008; Headden, online 2010 [wątpliwości metodologiczne budzą przesłanki 6 i 7 Headdena]). W | + | Kształt ozdoby jest niepewny. Tradycyjnie uznawano, że żagiel rozciągał się półkoliście nad grzbietem (Smith i in., 2006). Proponowano również, że najdłuższy wyrostek kolczysty znajdował się na ogonie (Cau, online 2008; Headden, online 2010 [wątpliwości metodologiczne budzą przesłanki 6 i 7 Headdena]). W rekonstrukcji Ibrahima i in. (2014), opartej również na FSAC-KK 11888, żagiel ma trapezoidalny kształt z dwoma wierzchołkami. Wysokość wyrostków kręgów krzyżowych i ogonowych jest w niej spekulatywna. Opisanie w 2020 r. niemal kompletnego ogona wykazało, że żagiel ciągnął się przez cały ogon, przyjmując formę podobną do tworów krokodyli bądź traszek. Wyrostki kolczyste przednich kręgów ogonowych były trzykrotnie wyższe od trzonów kręgów, a tylnych - aż siedmiokrotnie. Wydłużone były również szewrony, co zwiększało boczną powierzchnię ogona. Jednocześnie zredukowane były przed- i zazygapofyzy, co zwiększało giętkość ogona (Ibrahim i in., 2020b). |
| − | Stromer (1915) zauważył, że gładka powierzchnia i słaba konstrukcja górnych końców przeczy przypuszczeniu, by żagiel był pokryty silnymi mięśniami. Zaznaczył, że taki pogląd nie pasuje też do lokalizacji tej niezwykłej struktury | + | Stromer (1915) zauważył, że gładka powierzchnia i słaba konstrukcja górnych końców wyrostków przeczy przypuszczeniu, by żagiel był pokryty silnymi mięśniami. Zaznaczył, że taki pogląd nie pasuje też do lokalizacji tej niezwykłej struktury. Największe wyrostki kolczyste znajdowały się jego zdaniem w okolicy lędźwiowej, gdzie nie ma powodu do nadzwyczajnej rozbudowy mięśni grzbietu. Wg Stromera był to skórny żagiel, podobny do tego, który występuje u kameleona grzebieniastego (''Chamaeleo cristatus''). Obecne u obu gatunków pionowe prążki, ostre brzegi i duże odległości między wyrostkami również zdaniem Ibrahima i in. (2014) sugerują, że struktura spinozaura była pokryta warstwą skóry. |
Jack Bailey (1997) uznał, że żagiel nie był delikatną ozdobą pokrytą cienką warstwą skóry, a przedłużeniem grzbietu zwierzęcia, pokrytym mięśniami i ścięgnami. Jego zdaniem budowa wyrostków kolczystych kręgów spinozaura bardziej zbliża go do ssaków mających garb na grzbiecie, jak np. bizon, niż do bazalnych synapsydów, jak ''Dimetrodon''. U dinozaurów i ssaków wyrostki są masywniejsze i w widoku z góry wydłużone, a u bazalnych synapsydów są delikatne i okrągłe w przekroju. Wskazuje się jednak, że taka budowa u spinozaura jest po prostu dziedzictwem po przodkach (Holtz i in., 2004). | Jack Bailey (1997) uznał, że żagiel nie był delikatną ozdobą pokrytą cienką warstwą skóry, a przedłużeniem grzbietu zwierzęcia, pokrytym mięśniami i ścięgnami. Jego zdaniem budowa wyrostków kolczystych kręgów spinozaura bardziej zbliża go do ssaków mających garb na grzbiecie, jak np. bizon, niż do bazalnych synapsydów, jak ''Dimetrodon''. U dinozaurów i ssaków wyrostki są masywniejsze i w widoku z góry wydłużone, a u bazalnych synapsydów są delikatne i okrągłe w przekroju. Wskazuje się jednak, że taka budowa u spinozaura jest po prostu dziedzictwem po przodkach (Holtz i in., 2004). | ||
====Funkcje żagla==== | ====Funkcje żagla==== | ||
| − | Na temat funkcji powstało wiele hipotez, przy czym nowsze (od 2014 r.) często | + | Na temat funkcji tzw. "żagla" spinozaura powstało wiele hipotez, przy czym nowsze (od 2014 r.) często wiążą go z półwodnym trybem życia. |
| − | + | Żagiel mógł służyć do przyciągania samic i odstraszania skromniej obdarzonych rywali. Podobnie jak współczesne zwierzęta stosują swoje rogi, poroże lub kontrastową barwę podczas okresu godowego, również i spinozaur mógł używać swojego żagla by zainteresować wybredne samice. Samiec o większym i bardziej okazałym żaglu mógł wydawać się silniejszy lub zdrowszy. Możliwe także, że żagle w czasie zalotów napełniały się krwią w celu lepszej prezentacji. | |
| − | + | Inną możliwą funkcją żagla było powodowanie, że zwierzę wydawało się jeszcze większe, niż w rzeczywistości, co mogło mieć znaczenie w konkurowaniu z innymi mięsożercami. | |
| + | [[Plik:Spinotest con sigilmassa.jpg|300px|thumb|right|Rekonstrukcja szkieletu spinozaura zgodnie z koncepcją wysoko uniesionej szyi - Cau, online 2014 [http://theropoda.blogspot.it/2014/09/spinosaurus-revolution-episodio-iv-una.html].]] | ||
| + | Zdaniem Baileya (1997), struktura ta mogła służyć jako magazyn lipidów i substancji energetycznych (zwłaszcza przy dalekich wędrówkach lub w porze niedoboru pokarmu), pomagać w utrzymaniu gigantotermii lub stanowić osłonę przed gorącem. W związku z dużą masą takiej struktury, Bailey postulował czworonożność spinozaura. | ||
| − | + | [[Plik:Spinosaurus tidal harbinger tan by nashd1.jpg|300px|thumb|left|Rekonstrukcja spinozaura używającego swojego ciała do zablokowania ofiarom drogi ucieczki . Autor: Duane Nash [http://nashd1.deviantart.com/art/Tidal-Harbinger-tan-595706569].]] | |
| + | Cau (online 2016C), zaproponował, że u dwunożnego ''Spinosaurus'' wysokie wyrostki kolczyste kręgów były przystosowaniem do wydłużonego grzbietu. Cau posłużył się przykładem jednoprzęsłowego mostu budowanego z jednego brzegu kanionu, na którym umieszczony jest jedyny punkt podparcia. Wyjaśnił, że wraz z dokładaniem kolejnych przęseł (wydłużaniem mostu), podtrzymujące go liny muszą być przyczepione do coraz wyższego pylonu (wieży). W przypadku teropodów „linami” są mięśnie i ścięgna nadosiowe (''epaxial'') a pylonem wyrostki kolczyste. Wcześniej Cau (online 2013D) sugerował podobną funkcję na przykładzie koparek wielonaczyniowych. Cau (online 2014B), posługując się kręgami szyjnymi ''[[Sigilmassasaurus]]'', stwierdził, że ''Spinosaurus'' miał pionową, wysoko uniesioną szyję, niemal stykającą się z żaglem. Cau zasugerował, że ścięgna utrzymujące szyję w takim położeniu, bez użycia mięśni, były przyczepione do wyrostków kolczystych kręgów grzbietowych. Jest to problematyczna rekonstrukcja nie tylko z powodu użycia kręgów innego spinozauryda (''Spinosaurus'' mógł być przecież zbudowany podobnie), lecz także ze względu na podważenie rekonstrukcji „U-kształtnej” szyi ''Sigilmassasaurus'' Russella (1996) przez McFeetersa i in. (2013). Mortimer w komentarzach do postu Cau nie zgodził się z taką interpretacją a sam Cau niedostatecznie wyjaśnił swój pomysł. | ||
| − | + | Sugerowano, że żagiel mógł pełnić funkcję termoregulacyjną. Mógł być obficie unaczyniony i używany do nagrzania się w chłodniejsze, lecz nie zachmurzone dni i do ochładzania organizmu w czasie upałów. Mogło mieć to szczególne znaczenie przy ziemnowodnym trybie życia. Gdy dinozaur pływał w wodzie, żagiel zdaniem Ibrahima i in. (2014) wystawał nad jej powierzchnię. Wychwytywał zatem zapewne promienie słoneczne i ogrzewał zwierzę. Jeśli dinozaury, w tym ''Spinosaurus'', miały metabolizm pośredni między gadzim a ssaczym (Grady i in., 2014), to tego typu struktura mogła być niezwykle pomocna w środowisku wodnym, które szybko wychładza organizm. Wcześniej sugerowano, że hipoteza mezotermiczności wyjaśnia, dlaczego wśród dinozaurów nie było zwierząt wodnych (Sampson, 2009). Z pewnością również wielkie rozmiary pomagały spinozaurowi utrzymać ciepłotę ciała w wodzie. | |
| − | Ibrahim i in. (2014) sugerują, że funkcją pełnej w środku ozdoby było również dociążenie zwierzęcia | + | Ibrahim i in. (2014) sugerują, że funkcją pełnej w środku ozdoby było również dociążenie zwierzęcia. |
| − | + | Nash (online 2016) wskazuje, że żagiel mógł pomagać w łowieniu ryb przez zwiększanie powierzchni, którą spinozaur mógł blokować odpływ, uniemożliwiając ucieczkę ofiarom i je stłaczając. Żagiel mógł też służyć do zacieniania powierzchni wody, ułatwiając wypatrzenie zdobyczy. | |
| − | [[Plik:Spinosaurus & Sailfish comparison.png| | + | [[Plik:Spinosaurus & Sailfish comparison.png|200px|thumb|left|Rekonstrukcja oparta na hipotezie o hydrodynamicznej funkcji żagla. A - spinozaur w stanie spoczynku (szyja jest zbyt długa). B - współczesna żaglica. C- płynący pod wodą spinozaur. Żródło: Gimsa i in., 2016 [http://dx.doi.org/10.1017/S0016756815000801].]] |
| + | Inną hipotezę, związaną z podwodną lokomocją, przedstawili Gimsa i in. (2015). Ich zdaniem żagiel był strukturą analogiczną do płetwy grzbietowej żaglicy, której kształt odpowiada zrekonstruowanemu przez Ibrahima i in. (2014). Autorzy wykluczają wyłącznie pokazową funkcję żagla, gdyż byłby on przeszkodą na lądzie, a w wodzie musiałby przetrwać zanurzenie i zapewnić odpowiednią stabilność - w przeciwnym wypadku naraziłby zwierzę na uraz kręgosłupa. Ich zdaniem żagiel był używany przede wszystkim pod wodą i miał głównie hydrodynamiczne funkcje. Gimsa i in. sugerują, że żagiel był stabilizatorem – zapobiegał odchylaniu się tułowia podczas gwałtowniejszych ruchów głowy i szyi, co miałoby pozwalać na większy zasięg i szybkość ruchu łba w pogoni za zdobyczą (autorzy nie pokusili się jednak na dokładniejsze opisanie tego mechanizmu). Ponadto miałby działać podobnie jak miecz w łodzi, tj. zmniejszać znoszenie ciała na boki w wyniku bocznych ruchów ogona. Nash (online 2016) sugeruje, że żagiel pomagał zachowaniu równowagi (zapobiegał zataczaniu się) w skręcaniu i obracaniu się pod wodą. Hipoteza Gimsy i in. (2015) wydaje się prawdopodobna, choć wymaga dalszych badań. Pewne kwestie są jednak problematyczne - kształt żagla nie jest pewny a szyja została zrekonstruowana jako zbyt długa. | ||
===Postawa i lokomocja=== | ===Postawa i lokomocja=== | ||
| + | [[Plik:Spinosaurus quadruped by fredthedinosaurman.png|250px|thumb|right|Rekonstrukcja czworonożnego spinozaura. Autor: Fred Wierum [http://fredthedinosaurman.deviantart.com/art/The-COPROLITES-Dinosaur-Modes-494341269].]] | ||
| + | |||
====Poruszanie się na lądzie==== | ====Poruszanie się na lądzie==== | ||
| − | Opisanie FSAC-KK 11888 ożywiło pogląd o prawdopodobnej czworonożnej postawie ''Spinosaurus'' | + | Opisanie FSAC-KK 11888 ożywiło pogląd o prawdopodobnej czworonożnej postawie ''Spinosaurus''. Wcześniejsze hipotezy o czworonożności należy uznać za bezpodstawne, ponieważ nie były znane kości kończyn. |
| − | Typowa dla [[teropod]]ów dwunożna lokomocja była w przypadku ''Spinosaurus'' co najmniej znacznie utrudniona. Kończyny tylne były zmniejszone. Powierzchnia pełniącej funkcję przyczepu mięśni kości | + | Typowa dla [[teropod]]ów dwunożna lokomocja była w przypadku ''Spinosaurus'' co najmniej znacznie utrudniona. Kończyny tylne były zmniejszone. Powierzchnia pełniącej funkcję przyczepu mięśni kości biodrowej była około dwukrotnie mniejsza niż u innych teropodów. Grzebień nadpanewkowy, będący podporą kończyny, był niski. Dźwigająca ciężar ciała artykulacja stawu kolanowego była zredukowana (Ibrahim i in., 2014). Ponadto według rekonstrukcji z ww. publikacji, środek ciężkości był przesunięty do przodu. Zdaniem Ibrahima i in. (2014), uniemożliwiało to dwunożny chód, mogło jednak pomagać w utrzymaniu równowagi podczas pływania. |
| − | + | [[Plik:Spinosaurus Faster spino run run run toned by nashd1.jpg|300px|thumb|left|Rekonstrukcja uciekających na dwóch nogach młodych spinozaurów. Autor: Duane Nash [http://nashd1.deviantart.com/art/Faster-Spino-Run-Run-Run-Toned-595704097].]] | |
| − | Nash (online 2014A) zasugerował, że na lądzie ''Spinosaurus'' poruszał się prawdopodobnie przez ślizganie się na brzuchu. Wg | + | Wg Cau (online 2015) wysokie wyrostki kolczyste, smukły tors i budowa ogona sugerująca, że ''Spinosaurus'' mógł się nim podpierać, wskazują, że dinozaur mógł przyjmować półwyprostowaną, dwunożną postawę. W rekonstrukcji Cau użył również kręgów szyjnych ''[[Sigilmassasaurus]]'', które powodowały, że szyja była znacznie odgięta ku tyłowi, dodatkowo przesuwając środek ciężkości w tym kierunku (Cau, online 2014B). Wg Nasha (online 2016) młode okazy mogły biegać na dwóch nogach w półwyprostowanej pozycji. |
| + | |||
| + | Pełna czworonożność jest jednak mało prawdopodobna. Jedyny zilustrowany element kończyny przedniej znany u FSAC-KK 11888 (paliczek, kolejny element to część kolejnego paliczka) nie ma budowy sugerującej, że pełnił funkcje podporowe (Ibrahim i in., 2014, ryc. 2H). | ||
| + | |||
| + | Nash (online 2014A) zasugerował, że na lądzie ''Spinosaurus'' poruszał się prawdopodobnie przez ślizganie się na brzuchu. | ||
| + | |||
| + | Wg Hone i Holtza (2021) układ wyrostków bocznych ogona wskazuje na ułożenie mięśni kończyn tylnych typowe dla zwierząt lądowych. Nietypowa budowa stopy z szerokim rozstawem palców i użycie piątego palca w celach podporowych może świadczyć o przystosowaniu do poruszania się po lądzie, zwłaszcza po miękkim gruncie, i to pomimo zmniejszonych kończyn. Autorzy ci sugerują, że lokomocja lądowa spinozaura mogła się ograniczać do przemieszczania się między żerowiskami. | ||
| + | |||
| + | Wg Sereno i in. (2022) ''Spinosaurus'' na lądzie był zwierzęciem dwunożnym ze środkiem ciężkości umieszczonym nad stopami. Było to możliwe m.in. dzięki znacznej pneumatyzacji szkieletu osiowego w części przedkrzyżowej. Z kolej żagiel grzbietowy i ogon służyły głównie jako twory na pokaz (Sereno i in., 2022). | ||
====Poruszanie się w wodzie==== | ====Poruszanie się w wodzie==== | ||
Ibrahim i in. (2014) podali, że trzony kręgów ogonowych ''Spinosaurus'' były krótkie, a połączenia łuków neuralnych zredukowane, ułatwiając boczne zginanie ogona i wykorzystywanie go jako napędu. Dodatkowym napędem miałyby być stopy. Niepublikowane badania (Sereno i in., 2015) sugerują, że ''Spinosaurus'' był wolnym pływakiem. | Ibrahim i in. (2014) podali, że trzony kręgów ogonowych ''Spinosaurus'' były krótkie, a połączenia łuków neuralnych zredukowane, ułatwiając boczne zginanie ogona i wykorzystywanie go jako napędu. Dodatkowym napędem miałyby być stopy. Niepublikowane badania (Sereno i in., 2015) sugerują, że ''Spinosaurus'' był wolnym pływakiem. | ||
| − | Nash (online 2016) jako prawdopodobny sposób poruszania się wskazał nie pływanie, lecz chodzenie podwodne (dobijanie się od dna) na modłę hipopotamów. Wskazywać na to mają gęste kości, zmniejszona miednica i rozrost mięśnia ''caudofemoralis''. | + | [[Plik:Underwater runing spino page 001 by nashd1.jpg|300px|thumb|right|Rekonstrukcja podwodnego chodu spinozaura. Autor: Duane Nash [http://nashd1.deviantart.com/art/Underwater-Runing-Spino-page-001-569961605].]] |
| + | |||
| + | Nash (online 2016) jako prawdopodobny sposób poruszania się spinozaura w wodzie wskazał nie pływanie, lecz chodzenie podwodne (dobijanie się od dna) na modłę hipopotamów. Wskazywać na to mają gęste kości, zmniejszona miednica i rozrost mięśnia ''caudofemoralis''. | ||
| + | |||
| + | Poruszanie się w wodzie zapewne ułatwiała też specyficzna budowa kości czołowych, przez co oczy spinozaura były położone wyżej niż u innych teropodów i podniesione ponad resztę czaszki. Dzięki temu zwierzę mogło prawdopodobnie obserwować powierzchnię wody pomimo zanurzenia niemal całej czaszki (Arden i in. 2018). Z taką interpretacją nie zgodzili się Hone i Holtz (2021) którzy wykazali, że zachowanie możliwości obserwacji powierzchni wody wymagało wynurzenia niemal całej górnej części czaszki. Podobnie oddychanie nie było prawdopodobnie możliwe bez wystawiania niemal całej głowy, bowiem nozdrza spinozaura były wprawdzie cofniete do tyłu, ale nie skierowane do góry, jak u współczesnych krokodyli czy hipopotamów. Spinzaur bardziej przypominał w tym względzie ptaki brodzące, jak czaple. | ||
| + | |||
| + | W 2018 r. ukazała się praca Donalda Hendersona, podająca w wątpliwość półwodny tryb życia sinozaura. Autor posłużył się trójwymiarowym, cyfrowym modelem zwierzęcia, opartym w głównej mierze na rekonstrukcji z pracy Ibrahima i in. (2014). Został on porównany z modelem współczesnego aligatora oraz kilku innych teropodów, m.in. [[Suchomimus|zuchomima]] i [[Tyrannosaurus|tyranozaura]]. Badania wykazały, że środek ciężkości znajdował się znacznie bliżej bioder, niż sądzili Ibrahim i współpracownicy. Dzięki temu spinozaur wciąż byłby w pełni zdolny do poruszania się po lądzie. Autor stwierdził, że ''Spinosaurus'' z pewnością był w stanie unosić się na wodzie i oddychać z głową wystawioną nad jej powierzchnię, nie ma w tym jednak niczego specjalnego, gdyż większość zwierząt lądowych jest do tego przystosowana. Henderson wywnioskował, że spinozaur prawdopodobnie nie byłby w stanie całkowicie się zanurzyć z powodu zbyt małej średniej gęstości ciała, nawet po opróżnieniu płuc z powietrza w 75%. Stanowiłoby to bardzo dużą przeszkodę w ściganiu i chwytaniu wodnych zwierząt. Model skonstruowany przez Hendersona pokazał również, że utrzymanie pionowej postawy w wodzie byłoby bardzo trudne bez opierania się na kończynach, gdyż ciało spinozaura miałoby tendencję do przechylania się na bok. Wreszcie, podkreślił on niepewność co do poprawności rekonstrukcji w pracy Ibrahima i współpracowników. Wszystko to sprawia, że autor uznał spinozaura za teropoda wprawdzie mocno związanego z wodą, ale jego zdaniem nie było to wysoce wyspecjalizowane zwierzę, prowadzące półwodny tryb życia, ale raczej brodzące w płyciznach i aktywne również na lądzie. Do podobnych wniosków doszli Hone i Holtz (2021). Zauważyli oni, że silne wygięcie w dół szyi spinozaura, kształt czaszki, umiejscowienie nozdrzy i oczodołów predysponują go do polowania na linii brzegowej, z pyskiem częściowo zanurzonym lub znajdującym się tuż nad powierzchnią. Dzięki temu był on w stanie chwytać przepływające ryby za pomocą szybkich, pionowych ruchów głowy. Budowa zębów spinozaura wskazuje, że żywił się stosunkowo twardą wodną zdobyczą, taką jak ryby pancerne, skorupiaki i żółwie. | ||
| + | |||
| + | Ibrahim i in. (2020b) na podstawie opisu nowych szczątków podtrzymali swoje zdanie o spinozaurze jako zwierzęciu spędzającym większość czasu w wodzie. Głównym napędem byłby ogon, zbudowany inaczej niż u reszty teropodów. Dzięki zwiększonej powierzchni bocznej i dużej elastyczności, byłby on znacznie bardziej efektywnym napędem w wodzie niż ogony [[Coelophysis|celofyza]] i [[Allosaurus|allozaura]], z którymi go porównywano. Ustępował jednak w tym względzie ogonowi współczesnego krokodyla nilowego. | ||
| + | |||
| + | Hone i Holtz (2021) wykazali, że ogon spinozaura był stosunkowo sztywny u nasady a elastyczny na dalszym odcinku, co zdaje się podważać jego funkcje lokomotoryczne i wskazuje bardziej na funkcje związane z doborem płciowym. Autorzy ci uznali, że omawiany teropod był w stanie pływać, zapewne nawet lepiej niż inne teropody, ale raczej nie był zwierzęciem aktywnie polującym pod wodą. Takie zachowania byłyby także mocno utrudnione przez grzbietowy żagiel i stosunkowo duże kończyny przednie. Również badania Fisha i in. (2021) wykazały, że ogon spinozaura wytwarzał w wodzie znacznie mniejszy ciąg niż u współczesnego krokodyla. | ||
| + | |||
| + | Wg Beevora i współpracowników o wodnym trybie życia spinozaura świadczy też powszechne występowanie jego zębów w osadach rzecznych. Udział zębów typowo lądowych dinozaurów nie przekracza tam kilku procent, natomiast zęby spinozaura w niektórych miejscach są nawet liczniejsze niż zęby ryby ''Onchopristis''. Hone i Holtz (2021) uznali, że świadczy to o polowaniu spinozaura w rzekach (pod wodą lub na brzegu), ale nie przesądza o tym, czy spędzał on większość czasu zanurzony w wodzie. | ||
| + | |||
| + | Fabbri i in. (2022a) zaobserwowali u spinozaurydów wyraźny wzrost gęstości kości i m.in. na tej podstawie uznali ''Spinosaurus'' za nurkującego drapieżnika, zdolnego do polowania pod wodą. Wnioski z tej pracy były podważane przez Myhrvolda i in. (2022), którzy uznali dobór próbek za zbyt selektywny. Wskazywali też na nieścisłości dotyczące struktury kości spinozaurydów i błędne wnioski statystyczne. Ich zdaniem stosunkowo duża gęstość badanych próbek kości może wynikać po prostu ze znacznych rozmiarów ciała spinozaura, a nie przystosowań do nurkowania. Fabbri i in. (2022b) podtrzymali jednak swoje stanowisko w kolejnej publikacji. Myhrvold i in. (2024) ponownie skrytykowali wnioski Fabbriego i współpracowników. Podważyli m.in. możliwość wyciągania daleko idących wniosków z nielicznych i niepewnych szczątków. Ponadto zwrócili uwagę, że ze względu na wysoka pneumatyzację spinozaur prawdopodobnie nie był w stanie nurkować. Ich zdaniem ''Spinosaurus'' był półwodnym rybożercą, ale nie drapieżnikiem polującym aktywnie pod wodą. | ||
| + | |||
| + | Do podobnych wniosków doszli Sereno i in. (2022). Wg tych autorów, spinozaur był półwodnym drapieżnikiem polującym w strefie przybrzeżnej, ale nie zwierzęciem ściśle wodnym. Wykonany przez nich model cyfrowy ujawnił, że w głębokiej wodzie ''Spinosaurus'' byłby powolnym i niestabilnym pływakiem, niezdolnym do nurkowania z powodu zbyt dużej wyporności. Gęstość jego ciała oszacowano na 830 kg/m3, podczas gdy gęstość słonej wody wynosi 1026 kg/m3. | ||
| + | |||
| + | Wg Smarta i Sakamoto (2024) umiejscowienie nozdrzy spinozaura nie byłoby zbyt pomocne przy całkowicie wodnym trybie życia, jest natomiast użyteczne dla zwierząt takich jak rybożerne ptaki, brodzące w wodzie i polujące na przepływające ryby. | ||
| − | == | + | ===Wymiary=== |
| − | + | [[Plik:Spinosaurus_2_GetAwayTrike.png|300px|thumb|left|Rekonstrukcja holotypu i FSAC-KK 11888. Elementy zrekonstruowane mają kolor szary. Skala = 1 m. Autor: Genya Masukawa (GetAwayTrike) [http://blogs.yahoo.co.jp/rboz_05/36950355.html].]] | |
| − | [[Plik: | ||
| − | + | Już od odkrycia pierwszych kości w [[1912]] roku, ''Spinosaurus'' był jednym z najbardziej prawdopodobnych kandydatów do tytułu najdłuższego i najcięższego [[teropod]]a. Wszelkie oszacowania sprzed opisania FSAC-KK 11888 w 2014 r. należy jednak pominąć, jako opartych na nieprawidłowych rekonstrukcjach. | |
| − | Już od odkrycia pierwszych kości w [[1912]] roku, ''Spinosaurus'' był jednym z najbardziej prawdopodobnych kandydatów do tytułu najdłuższego i najcięższego [[teropod]]a. Wszelkie oszacowania sprzed opisania FSAC-KK 11888 w 2014 r. należy pominąć. | ||
Wg Ibrahima i in. (2014) maksymalne rozmiary dorosłego osobnika to 15 m długości. Oszacowanie masy według ich rekonstrukcji wykazało wartość 7.557 kg (Franoys, online 2016); przy identycznych założeniach jak Hartmana (online 2013), który uzyskał dla ''[[Tyrannosaurus]]'' FMNH PR2081 8.400 kg i dla ''[[Giganotosaurus]]'' MUCPv-95 8.200 kg. Zwiększenie gęstości kończyn i żagla o hipotetyczne i zapewne zawyżone 20% (por. np. Wedel, 2007; Henderson, 2004) powoduje, że masa zwiększa się o ok. 154 kg - do 7.713 t. | Wg Ibrahima i in. (2014) maksymalne rozmiary dorosłego osobnika to 15 m długości. Oszacowanie masy według ich rekonstrukcji wykazało wartość 7.557 kg (Franoys, online 2016); przy identycznych założeniach jak Hartmana (online 2013), który uzyskał dla ''[[Tyrannosaurus]]'' FMNH PR2081 8.400 kg i dla ''[[Giganotosaurus]]'' MUCPv-95 8.200 kg. Zwiększenie gęstości kończyn i żagla o hipotetyczne i zapewne zawyżone 20% (por. np. Wedel, 2007; Henderson, 2004) powoduje, że masa zwiększa się o ok. 154 kg - do 7.713 t. | ||
| − | Mortimer (online 2016) podaje, że [[holotyp]] mierzył 13,2 m | + | Mortimer (online 2016) podaje, że [[holotyp]] mierzył 13,2 m przy długości czaszki 1,45 m. Wg Cau (online 2014C) okaz mierzył 12-13 m a czaszka mogła mieć ok. 1,32 m (Cau, online 2013C; okaz MSNM V4047 Cau szacuje na ok. 1,3 m i czasem na 1,4 m i uważa również, że jest on 5-10% większy od holotypu – zob. [[Spinosauridae#Wymiary]]). Cau twierdził także (online 2013A), że okaz jest porównywalnych rozmiarów z holotypem ''[[Baryonyx]]''. Wg rekonstrukcji obok długość ciała to ok. 11 m a czaszki - 1,15-1,25 m. Zdania na temat dojrzałości okazu są podzielone. Niektórzy podają (np. Paul, 1988), że był to okaz niedorosły ze względu na niezrośnięte trzony kręgów grzbietowych i ich wyrostki. Inni twierdzą (np. Cau, online 2013C), że okaz jest dorosły. Stromer (1915) wskazał, że w kręgu szyjnym trzon jest „połączony z wyrostkiem neuralnym szwem” a na ilustracjach trzony kręgów grzbietowych i ich wyrostki są w większości dysartykułowane (wyjątkiem jest kręg „f”). Wyraźnie widoczne są jednak szwy między kręgami krzyżowymi, których obecność u [[bazalny]]ch (nie[[celurozaur]]owych) [[tetanur]]ów sugeruje niedojrzałość (Brusatte i in., 2008, s.33; Rauhut, 2005). |
| + | [[Plik:Spinosaurus_2020_reconstruction.jpg|300px|thumb|left|Rekonstrukcja uwzględniająca najnowsze informacje o ogonie, znacznie wyższym niż wcześniej sądzono. Autor: Mario Lanzas [https://commons.wikimedia.org/wiki/File:Spinosaurus_2020_reconstruction.jpg].]] | ||
| − | Ibrahim i in. (2014) szacują, że FSAC-KK 11888 - ok. 17-letni, nie w pełni wyrośnięty osobnik - był o 32% mniejszy niż największe okazy (tj. 15-metrowe), miałby więc ok. 10,2 m. Nie zgadza się to jednak z danymi podanymi w suplemencie. W tab. S1 podano, że długość od końca pyska do tylnego końca miednicy wynosi | + | Ibrahim i in. (2014) szacują, że FSAC-KK 11888 - ok. 17-letni, nie w pełni wyrośnięty osobnik - był o 32% mniejszy niż największe okazy (tj. 15-metrowe), miałby więc ok. 10,2 m. Nie zgadza się to jednak z danymi podanymi w suplemencie. W tab. S1 podano, że długość od końca pyska do tylnego końca miednicy wynosi u niego 5,88 m, więc na ogon pozostałoby 4,32 m (42% całkowitej długości). Jednak na rekonstrukcji szkieletu (Ibrahim i in., 2014, rys. F3) ogon stanowi ok. 48% długości ciała. Ibrahim (prywatna korespondencja do MZ, 2015) wyjaśnia, że rekonstrukcja nie jest „płaska”, więc mogą występować kilkuprocentowe różnice. Mortimer (online 2016) długość tego okazu oszacował na ok. 11,3 m, Cau (online 2014C) na 11 m a wg rekonstrukcji obok ma ok. 9,5 m. |
| − | BSPG 2006 I 57 – szósty kręg szyjny – ma długość 19,8 cm (Evers i in., 2015), jest więc ok. 7% dłuższy niż piąty kręg szyjny holotypu (18,5 cm - Stromer, 1915). Jest od niego także szerszy (9,1 do ok. 7 cm), lecz niższy (8,5 do ok. 10 cm [w przedniej strony trzonu] oraz 10 do ok. 12,5 cm [w tylnej części trzonu | + | BSPG 2006 I 57 – szósty kręg szyjny (piąty wg Smytha i in., 2020) – ma długość 19,8 cm (Evers i in., 2015), jest więc ok. 7% dłuższy niż piąty kręg szyjny holotypu (18,5 cm - Stromer, 1915). Jest od niego także szerszy (9,1 do ok. 7 cm), lecz niższy (8,5 do ok. 10 cm [w przedniej strony trzonu] oraz 10 do ok. 12,5 cm [w tylnej części trzonu; wysokość u holotypu oszacowana według ryc. 2 z tabl. 2 z pracy Stromera]), co potwierdza, że kręg okazu typowego jest bocznie zgnieciony (Stromer, 1915). Trudno zatem ustalić, który z okazów reprezentuje większego osobnika. Przyjęcie, że BSPG 2006 I 57 reprezentuje okaz większy o 7% od typowego wskazuje na jego długość w granicach 11,8-14,1 m. |
| + | |||
| + | Ibrahim i in. (2020b) wykorzystując najnowsze znaleziska i dodatkowy materiał okazu FSAC-KK 11888, oszacowali jego masę na 3,2-4,2 t, wykorzystując różne warianty gęstości tkanek. Największe osobniki ważyłyby zatem 5,4-7,1 t. | ||
| + | |||
| + | ==Środowisko== | ||
| + | [[Plik:Spinosaurus with contemporaneous taxa.jpg|300px|thumb|left|''Spinosaurus'' w towarzystwie współczesnych mu zwierząt wodnych. Autor: Joschua Knüppe. Źródło: Jamale Ijouiher , 2016 [https://peerj.com/preprints/2470/].]] | ||
| − | + | Spinozaury, zamieszkujące Afrykę Północną na początku późnej kredy, mogły żyć w pobliżu lasów namorzynowych, polując w zbiornikach wodnych na gady lub ryby. Obok tego wielkiego drapieżcy żyło szczególnie wiele teropodów - zob. [[Carcharodontosaurus#Środowisko]] i [[Spinosauridae#Paleobiologia]]. | |
| − | |||
==Spinozaur w kulturze masowej== | ==Spinozaur w kulturze masowej== | ||
| − | + | [[Plik:Spinosaurus_nyansaurus_aegyptiacus_by_osmatar.jpg|100px|thumb|left|”Nyansaurus aegyptiacus”. Autor: Osmatar [http://osmatar.deviantart.com/art/Nyansaurus-aegyptiacus-485897880].]] | |
| + | [[Plik:Spinosaurus_Spinozilla_by_fredthedinosaurman.jpg|200px|thumb|right|”Spinozilla”. Autor: Fred Wierum [http://fredthedinosaurman.deviantart.com/art/SpinoZilla-The-perfect-blend-between-507512527].]] | ||
| + | ''Spinosaurus'' jest dobrze znany przez wielbicieli dinozaurów z powodu wielkości, nietypowego żagla i wydłużonej krokodylej czaszki. | ||
| + | |||
| + | Spinozaur od dawna przedstawiany był w książkach popularnonaukowych o dinozaurach, lecz dopiero niedawno pojawiły się dokładniejsze informacje umożliwiające pełniejszy opis zwierzęcia. Od rekonstrukcji [[Albert-Félix Lapparent|Lapparenta]] i Lavocata z [[1955]] roku, spinozaura wyobrażano sobie jako dużego teropoda z wysoką, silną czaszką (jak u innych wielkich teropodów) i charakterystycznym grzbietowym żaglem, czasami w pozycji czworonożnej. Później, w związku z nowymi odkryciami, zrewidowano kształt łba. Zarówno starsze, jak i wiele nowych wizerunków spinozaura często nie oddająle jego rzeczywistej budowy, zniekształcając np. kształt żuchwy czy wyrostków kręgów, które zilustrowano ponad sto lat temu. | ||
| + | |||
| + | Spinozaur wystąpił w filmie Park Jurajski III z [[2001]] roku, w którym to przedstawiono go jako większego i silniejszego niż ''[[Tyrannosaurus]]''. W jednej z scen spinozaur po długiej walce pokonuje tyranozaura, wykręcając mu kark. W rzeczywistości tego typu pojedynek nie mógł się wydarzyć, ponieważ teropody te dzieliło nie tylko odległe miejsce występowania ale i kilkadziesiąt milionów lat. Scena ta wywołała niemałe kontrowersje wśród fanów tyranozaura. | ||
| + | |||
| + | "Kolczasty jaszczur" wystąpił również w grach Jurassic Park: Operation Genesis i Zoo Tycoon: Dinosaur Digs, w których także był dominującym drapieżcą, | ||
| − | Oprócz tego ''Spinosaurus'' został przedstawiony w kilku programach popularnonaukowych. W produkcji Mega Beasts (polski tytuł Prehistoryczne Bestie), wyprodukowanym przez Discovery Channel w 2009 r. poświęcono mu w całości jeden z odcinków, zatytułowany Biggest Killer Dino. Ukazano tam spinozaura jako czołowego drapieżcę swoich czasów i niekwestionowanego władcę jego świata, z którym nie mógł się mierzyć ''[[Carcharodontosaurus]]'' ani żadne inne zwierzę. Bardziej stonowany wizerunek omawianego teropoda przedstawiono w serialu BBC Planet Dinosaur z 2011 r. Odcinek zatytułowany New Giants przedstawia Północną Afrykę sprzed 95 mln lat. ''Spinosaurus'' został tam ukazany jako wyspecjalizowany rybożerca, unikający konkurencji z karcharodontozaurem dzięki zajmowaniu odmiennej niszy ekologicznej. Mimo to w warunkach braku pożywienia stacza z nim zaciętą, ale zwycięską walkę, okupioną jednak poważnymi ranami. Kolejnym programem dokumentalnym | + | Oprócz tego ''Spinosaurus'' został przedstawiony w kilku programach popularnonaukowych. W produkcji Mega Beasts (polski tytuł Prehistoryczne Bestie), wyprodukowanym przez Discovery Channel w 2009 r. poświęcono mu w całości jeden z odcinków, zatytułowany Biggest Killer Dino. Ukazano tam spinozaura jako czołowego drapieżcę swoich czasów i niekwestionowanego władcę jego świata, z którym nie mógł się mierzyć ''[[Carcharodontosaurus]]'' ani żadne inne zwierzę. Bardziej stonowany wizerunek omawianego teropoda przedstawiono w serialu BBC Planet Dinosaur z 2011 r. Odcinek zatytułowany New Giants przedstawia Północną Afrykę sprzed 95 mln lat. ''Spinosaurus'' został tam ukazany jako wyspecjalizowany rybożerca, unikający konkurencji z karcharodontozaurem dzięki zajmowaniu odmiennej niszy ekologicznej. Mimo to w warunkach braku pożywienia stacza z nim zaciętą, ale zwycięską walkę, okupioną jednak poważnymi ranami. Kolejnym programem dokumentalnym poświęconym spinozaurowi była produkcja zatytułowana „Bestia”. Opowiada ona o przełomowym odkryciu Ibrahima i współpracowników. Po raz pierwszy zaprezentowano obraz wodnego, czworonożnego rybożercy. |
| + | |||
| + | Oprócz rekonstrukcji, starających się wiarygodnie odtworzyć wygląd, środowisko i zachowanie zwierzęcia, spinozaur jest także tematem prac żartobliwych, jak np. odnoszących się do popularnych memów internetowych czy Godzilli. | ||
==Spis gatunków== | ==Spis gatunków== | ||
| Linia 254: | Linia 316: | ||
| {{V|''S. aegyptiacus''}} | | {{V|''S. aegyptiacus''}} | ||
| {{Kpt|[[Ernst Stromer|Stromer]]}}, [[1915]] | | {{Kpt|[[Ernst Stromer|Stromer]]}}, [[1915]] | ||
| − | |||
|- | |- | ||
| ''S. maroccanus'' | | ''S. maroccanus'' | ||
| Linia 260: | Linia 321: | ||
| = ''[[Sigilmassasaurus]] brevicollis'' | | = ''[[Sigilmassasaurus]] brevicollis'' | ||
|} | |} | ||
| + | |||
| + | ==Galeria== | ||
| + | <gallery> | ||
| + | Plik:Spinosaurus_by_Joschua_Knüppe.png|Rekonstrukcja. Autor: Joschua Knüppe [https://peerj.com/preprints/2470]. | ||
| + | |||
| + | Plik:Spinosaurus aegyptiacus by abelov2014-d8m78hv.jpg|Rekonstrukcja (na drugim planie niepoprawnie przedstawiono roślinożerne ''[[Ouranosaurus]]'', które żyły na tych terenach wcześniej). Autor: ABelov2014 [http://abelov2014.deviantart.com/art/Spinosaurus-aegyptiacus-521018563]. | ||
| + | |||
| + | Plik:Spinosaurus Chunky spino by arvalis.jpg|Rekonstrukcja. Autor: RJ Palmer (arvalis) [http://arvalis.deviantart.com/art/Chunky-Spino-482388387]. | ||
| + | |||
| + | Plik:Спинозавр - новая реконструкция.jpg|Rekonstrukcja płynącego spinozaura. Autor: ДиБгд. [https://commons.wikimedia.org/wiki/File:%D0%A1%D0%BF%D0%B8%D0%BD%D0%BE%D0%B7%D0%B0%D0%B2%D1%80_-_%D0%BD%D0%BE%D0%B2%D0%B0%D1%8F_%D1%80%D0%B5%D0%BA%D0%BE%D0%BD%D1%81%D1%82%D1%80%D1%83%D0%BA%D1%86%D0%B8%D1%8F.jpg]. | ||
| + | |||
| + | Plik:Spinosaurus_durbed.jpg|Rekonstrukcja. Autor: Durbed [http://durbed.deviantart.com/art/Spinosaurus-Aegyptiacus-287547465]. | ||
| + | |||
| + | Plik:Spinosaurus by tomozaurus.png|Rekonstrukcja sprzed 2014 r. Autor: Tom Parker [http://tomozaurus.deviantart.com/art/Spinosaurus-447883270]. | ||
| + | Plik:Spinosaurus_white_background_2.jpg|Rekonstrukcja szkieletu. Fot.: Mariomassone [https://commons.wikimedia.org/wiki/File:Spinosaurus_white_background_2.jpg] | ||
| + | |||
| + | |||
| + | Plik:Spinosaurus_attacking_Onchopristis.jpg|Rekonstrukcja szkieletu. Fot.: Mike Bowler [https://www.flickr.com/photos/mbowler/15116295389/] | ||
| + | |||
| + | Plik:Spinosaurus_back_view.jpg|Rekonstrukcja szkieletu. Fot.: Ryan Somma [https://www.flickr.com/photos/ideonexus/16065918552] | ||
| + | |||
| + | Plik:Spinosaurus_skull_steveoc.jpg|Rekonstrukcja głowy. Autor: Stephen O'Connor [https://commons.wikimedia.org/wiki/File:Spinosaurus_skull_steveoc.jpg] | ||
| + | |||
| + | Plik:Spinosaurus by fredthedinosaurman.jpg|Rekonstrukcja łba, oparta na wyglądzie i zachowaniu żółwi jaszczurowatych. Autor: Fred Wiernum [http://fredthedinosaurman.deviantart.com/art/Snapping-Spinosaurus-491342697]. | ||
| + | |||
| + | Plik:Spinosaurus_new_skull.jpg|Rekonstrukcja czaszki. Fot.: Mike Bowler [https://www.flickr.com/photos/mbowler/15116294799/] | ||
| + | |||
| + | Plik:3D_printed_Spinosaurus_skulls.jpg|Rekonstrukcja czaszki. Fot.: Ryan Somma [https://www.flickr.com/photos/ideonexus/1604083811] | ||
| + | |||
| + | Plik:Spinosaurus_muscles.jpg|Rekonstrukcja czaszki i muskulatury. Autor: Christophe Hendrickx [https://commons.wikimedia.org/wiki/File:Spinosaurus_muscles.jpg] | ||
| + | |||
| + | Plik:Spinosaurus_skull_en.svg|Rekonstrukcja czaszki. Autor: AS [https://commons.wikimedia.org/wiki/File:Spinosaurus_skull_en.svg] | ||
| + | |||
| + | Plik:Fig-18-full.png|Kręg szyjny (C?4) NHMUK PV R 16429. Źródło: Evers i in., 2015 [http://dx.doi.org/10.7717/peerj.1323]. | ||
| + | |||
| + | Plik:Fig-21-full.png|Kręg szyjny (C6-7) BSPG 2012 I 97. Źródło: Evers i in., 2015 [http://dx.doi.org/10.7717/peerj.1323]. | ||
| + | |||
| + | Plik:Spinosaurus_foot_restoration.JPG|Rekonstrukcja stopy. Fot.: Ghedoghedo [https://commons.wikimedia.org/wiki/File:Spinosaurus_foot_restoration.JPG] | ||
| + | |||
| + | </gallery> | ||
==Bibliografia== | ==Bibliografia== | ||
| − | <small>Amiot, R., Buffetaut, E., Lécuyer, C., Wang, X., Boudad, L., Ding, Z., Fourel, F., Hutt, S., Martineau, F., Medeiros, M.A., Mo, J., Simon, L., Suteethorn, V., Sweetman, S., Tong, H., Zhang, F. & Zhou, Z. (2010) "Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods" Geology 38, 139-142. | + | <small>Arden, T.M.S., Klein, C.G., Zouhri, S. & Longrich, N.R. (2018). “Aquatic adaptation in the skull of carnivorous dinosaurs (Theropoda: Spinosauridae) and the evolution of aquatic habits in spinosaurus.” Cretaceous Research. [[doi: https://doi.org/10.1016/j.cretres.2018.06.013]]. |
| + | |||
| + | Amiot, R., Buffetaut, E., Lécuyer, C., Wang, X., Boudad, L., Ding, Z., Fourel, F., Hutt, S., Martineau, F., Medeiros, M.A., Mo, J., Simon, L., Suteethorn, V., Sweetman, S., Tong, H., Zhang, F. & Zhou, Z. (2010) "Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods" Geology 38, 139-142. | ||
Bailey, J.B. (1997) "Neural spine elongation in dinosaurs: Sailbacks or buffalo-backs?" Journal of Paleontology, 71(06), 1124-1146. | Bailey, J.B. (1997) "Neural spine elongation in dinosaurs: Sailbacks or buffalo-backs?" Journal of Paleontology, 71(06), 1124-1146. | ||
| + | |||
| + | Beevor, T., Quigley, A., Smith, R.E., Smyth, R.S.H., Ibrahim, N., Zouhri, S. & Martill, D.M. (2020). "Taphonomic evidence supports an aquatic lifestyle for ''Spinosaurus''." Cretaceous Research 104627 (advance online publication) | ||
| + | [[doi: https://doi.org/10.1016/j.cretres.2020.104627]]. | ||
Brusatte, S.L., Benson, R.B.J. & Hutt, S. (2008) "The osteology of Neovenator salerii (Dinosauria: Theropoda) from the Wealden (Barremian) of the Isle ofWight" Monograph of the Palaeontographical Society, 162(631), 1-166. | Brusatte, S.L., Benson, R.B.J. & Hutt, S. (2008) "The osteology of Neovenator salerii (Dinosauria: Theropoda) from the Wealden (Barremian) of the Isle ofWight" Monograph of the Palaeontographical Society, 162(631), 1-166. | ||
| Linia 301: | Linia 407: | ||
Evers, S.W., Rauhut, O.W.M., Milner, A.C., McFeeters, B. & Allain, R. (2015) "A reappraisal of the morphology and systematic position of the theropod dinosaur Sigilmassasaurus from the “middle” Cretaceous of Morocco" PeerJ, 3, e1323. [[doi:10.7717/peerj.1323]] | Evers, S.W., Rauhut, O.W.M., Milner, A.C., McFeeters, B. & Allain, R. (2015) "A reappraisal of the morphology and systematic position of the theropod dinosaur Sigilmassasaurus from the “middle” Cretaceous of Morocco" PeerJ, 3, e1323. [[doi:10.7717/peerj.1323]] | ||
| + | |||
| + | Fabbri, M., Navalón, G., Benson, R.B.J., Pol, D., O’Connor, J., Bhullar, B.-A.S., Erickson, G.M., Norell, M.A., Orkney, A., Lamanna, M.C., Zouhri, S., Becker, J., Emke, A., Dal Sasso, C., Bindellini, G., Maganuco, S., Auditore, M. & Ibrahim, N. (2022a) "Subaqueous foraging among carnivorous dinosaurs." Nature 603, 852–857 (2022). [[https://doi.org/10.1038/s41586-022-04528-0]]. | ||
| + | |||
| + | Fabbri, M., Navalón, G., Benson, R.B.J., Pol, D., O’Connor, J., Bhullar, B.-A.S., Erickson, G.M., Norell, M.A., Orkney, A., Lamanna, M.C., Zouhri, S., Becker, J., Emke, A., Dal Sasso, C., Bindellini, G., Maganuco, S., Auditore, M. & Ibrahim, N. (2022b) "Sinking a giant: quantitative macroevolutionary comparative methods debunk qualitative assumptions." bioRxiv, [[https://doi.org/10.1101/2022.05.05.490811]]. | ||
| + | |||
| + | Fish, F.E., Rybczynski, N., Lauder, G.V., & Duff, C.M. (2021) "The role of the tail or lack thereof in the evolution of tetrapod aquatic propulsion." Integrative and Comparative Biology, 61:398-413. [[https://doi.org/10.1093/icb/icab021]] | ||
Franoys (online 2016) http://franoys.deviantart.com/journal/A-mathematical-analysis-on-Spinosaurus-mass-635495737 | Franoys (online 2016) http://franoys.deviantart.com/journal/A-mathematical-analysis-on-Spinosaurus-mass-635495737 | ||
| Linia 315: | Linia 427: | ||
Headden, J. (online 2014) https://qilong.wordpress.com/2014/09/12/the-outlaw-spino-saurus/ | Headden, J. (online 2014) https://qilong.wordpress.com/2014/09/12/the-outlaw-spino-saurus/ | ||
| + | |||
| + | Henderson, D.M. (2018). “A buoyancy, balance and stability challenge to the hypothesis of a semi-aquatic Spinosaurus Stromer, 1915 (Dinosauria: Theropoda).” PeerJ 6:e5409Â. [[https://doi.org/10.7717/peerj.5409]]. | ||
Hendrickx, C., Mateus, O. & Buffetaut, E. (2016) "Morphofunctional analysis of the quadrate of Spinosauridae (Dinosauria: Theropoda) and the presence of Spinosaurus and a second Spinosaurine taxon in the Cenomanian of North Africa" PLoS ONE, 11(1), e0144695. [[doi:10.1371/journal.pone.0144695]] | Hendrickx, C., Mateus, O. & Buffetaut, E. (2016) "Morphofunctional analysis of the quadrate of Spinosauridae (Dinosauria: Theropoda) and the presence of Spinosaurus and a second Spinosaurine taxon in the Cenomanian of North Africa" PLoS ONE, 11(1), e0144695. [[doi:10.1371/journal.pone.0144695]] | ||
| + | |||
| + | Hone, D.W.E. & Holtz, T.R.Jr. (2021) “Evaluating the ecology of Spinosaurus: Shoreline generalist or aquatic pursuit specialist?” Palaeontologia Electronica 24(1):a03 | ||
| + | [[doi: https://doi.org/10.26879/1110]] | ||
Holtz, T.R., Molnar, R.E. & Currie, P.J. (2004) "Basal Tetanurae" [w:] Weishampel, D.B., Dodson, P. & Osmólska, H. "The Dinosauria" Berkeley and Los Angeles: University of California Press, 71-110. | Holtz, T.R., Molnar, R.E. & Currie, P.J. (2004) "Basal Tetanurae" [w:] Weishampel, D.B., Dodson, P. & Osmólska, H. "The Dinosauria" Berkeley and Los Angeles: University of California Press, 71-110. | ||
| − | Ibrahim, N., Sereno, P.C., Dal Sasso, C., Maganuco, S., Fabbri, M., Martill, D.M., Zouhri, S., Myhrvold, N. & Iurino, D.A. (2014) "Semiaquatic adaptations in a giant predatory dinosaur" Science, 345(6204), 1613-1616. doi:10.1126/science.1258750 | + | Ibrahim, N., Sereno, P.C., Dal Sasso, C., Maganuco, S., Fabbri, M., Martill, D.M., Zouhri, S., Myhrvold, N. & Iurino, D.A. (2014) "Semiaquatic adaptations in a giant predatory dinosaur" Science, 345(6204), 1613-1616. [[doi:10.1126/science.1258750]]. |
| + | |||
| + | Ibrahim, N., Sereno, P.C., Varricchio, D.J., Martill, D.M., Dutheil, D.B., Unwin, D.W., Baidder, L., Larsson, H.C.F., Zouhri, S. & Kaoukaya, A/ (2020a). "Geology and paleontology of the Upper Cretaceous Kem Kem Group of eastern Morocco". ZooKeys. 928: 1–216. [[doi:10.3897/zookeys.928.47517]]. | ||
| + | |||
| + | Ibrahim, N., Maganuco, S., Dal Sasso, C., Fabbri, M., Auditore, M., Bindellini, G., Martill, D.M., Zouhri, S., Mattarelli, D.A., Unwin, D.M., Wiemann, J., Bonadonna, D., Amane, A., Jakubczak, J., Joger, U., Lauder, G.V & Pierce, S.E. (2020b). "Tail-propelled aquatic locomotion in a theropod dinosaur." Nature (2020). [[https://doi.org/10.1038/s41586-020-2190-3]]. | ||
| + | |||
| + | Lacerda, M.B.S., Grillo, O.N. & Romano, P.S.R. (2021): Rostral morphology of Spinosauridae (Theropoda, Megalosauroidea): premaxilla shape variation and a new phylogenetic inference, Historical Biology, [[DOI:10.1080/08912963.2021.2000974]]. | ||
| + | |||
| + | Lakin, R.J & Longrich, N.R. (2018) “Juvenile spinosaurs (Theropoda: Spinosauridae) from the middle Cretaceous of Morocco and implications for spinosaur ecology.” Cretaceous Research (in press). [https://www.sciencedirect.com/science/article/pii/S0195667118302052] | ||
| + | |||
| + | Maganuco, S. & Dal Sasso, C. (2018. "The smallest biggest theropod dinosaur: a tiny pedal ungual of a juvenile Spinosaurus from the Cretaceous of Morocco". PeerJ 6:e4785. [[doi: https://doi.org/10.7717/peerj.4785]]. | ||
McFeeters, B., Ryan, M.J., Hinic-Frlog, S., Schröder-Adams, C. & Sues, H. (2013) "A reevaluation of Sigilmassasaurus brevicollis (Dinosauria) from the Cretaceous of Morocco" Canadian Journal of Earth Sciences, 50(6), 636-649. [[doi:10.1139/cjes-2012-0129]] | McFeeters, B., Ryan, M.J., Hinic-Frlog, S., Schröder-Adams, C. & Sues, H. (2013) "A reevaluation of Sigilmassasaurus brevicollis (Dinosauria) from the Cretaceous of Morocco" Canadian Journal of Earth Sciences, 50(6), 636-649. [[doi:10.1139/cjes-2012-0129]] | ||
| Linia 327: | Linia 454: | ||
Mortimer, M. (online 2016) http://theropoddatabase.com/Megalosauroidea.htm#Spinosaurusaegyptiacus | Mortimer, M. (online 2016) http://theropoddatabase.com/Megalosauroidea.htm#Spinosaurusaegyptiacus | ||
| + | |||
| + | Myhrvold, N., Sereno, P.C., Baumgart, S.L., Formoso, K.K., Vidal, D., Fish, F.E. & Henderson, D.E. (2022) "Spinosaurids as “subaqueous foragers” undermined by selective sampling and problematic statistical inference.". bioRxiv [[doi: https://doi.org/10.1101/2022.04.13.487781]] | ||
| + | |||
| + | Myhrvold, N.P., Baumgart. S.L., Vidal, D., Fish, F.E., Henderson, D.M. & Saitta, E.T. (2024) "Diving dinosaurs? Caveats on the use of bone compactness and pFDA for inferring lifestyle." PLoS ONE 19(3): e0298957. [[https://doi.org/10.1371/journal.pone.0298957]] | ||
Nash, D. (online 2014A) http://antediluviansalad.blogspot.com/2014/08/did-bakker-get-spinosaurus-right-after.html | Nash, D. (online 2014A) http://antediluviansalad.blogspot.com/2014/08/did-bakker-get-spinosaurus-right-after.html | ||
| Linia 348: | Linia 479: | ||
Sereno, P.C., Beck, A.L., Dutheil, D.B., Gado, B., Larsson, H.C.E., Lyon, G.H., Marcot, J.D., Rauhut, O.W.M., Sadleir, R.W., Sidor, C.A., Varrichio, D.D., Wilson, G.P. & Wilson, J.A. (1998) "A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids" Science 282, 1298-1302. [[doi:10.1126/science.282.5392.1298]] | Sereno, P.C., Beck, A.L., Dutheil, D.B., Gado, B., Larsson, H.C.E., Lyon, G.H., Marcot, J.D., Rauhut, O.W.M., Sadleir, R.W., Sidor, C.A., Varrichio, D.D., Wilson, G.P. & Wilson, J.A. (1998) "A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids" Science 282, 1298-1302. [[doi:10.1126/science.282.5392.1298]] | ||
| − | Sereno, P.C., Fish, F.E. & Myhrvold, N. (2015) " Swimming function in the Cretaceous giant Spinosaurus aegyptiacus based on the kinematics of undulatory swimming in the American alligator" Journal of Vertebrate Paleontology, Program and Abstracts, 153. | + | Sereno, P.C., Fish, F.E. & Myhrvold, N. (2015) "Swimming function in the Cretaceous giant Spinosaurus aegyptiacus based on the kinematics of undulatory swimming in the American alligator" Journal of Vertebrate Paleontology, Program and Abstracts, 153. |
| + | |||
| + | Sereno, P.C., Myhrvold, N., Henderson, D.M., Fish, F.E., Vidal, D., Baumgart, S.L., Keillor, T.M., Formoso, K.K. & Conroy, L.L. (2022). "Spinosaurus is not an aquatic dinosaur.". bioRxiv (preprint). | ||
| + | [[doi: https://doi.org/10.1101/2022.05.25.493395]] | ||
| + | |||
| + | Smart, S. & Sakamoto, M. (2024). "Using linear measurements to diagnose the ecological habitat of ''Spinosaurus''. PeerJ 12:e17544 [[DOI 10.7717/peerj.17544]]. | ||
Smith, J.B., Lamanna, M.C., Mayr, H., & Lacovara, K.J. (2006) "New information regarding the holotype of Spinosaurus aegyptiacus Stromer, 1915" Journal of Paleontology, 80(2), 400-406. | Smith, J.B., Lamanna, M.C., Mayr, H., & Lacovara, K.J. (2006) "New information regarding the holotype of Spinosaurus aegyptiacus Stromer, 1915" Journal of Paleontology, 80(2), 400-406. | ||
| + | |||
| + | Smith, R.E. & Martill, D.W. (2023). "An unusual dental pathology in a tooth of ''Spinosaurus'' (Dinosauria, Theropoda) from the mid-Cretaceous of Morocco. Cretaceous Research. 146. 105499, [[https://doi.org/10.1016/j.cretres.2023.105499]]. | ||
| + | |||
| + | Smyth, R.S.H., Ibrahim, N. & Martill, D.M. (2020)/ "''Sigilmassasaurus'' is ''Spinosaurus'': a reappraisal of African spinosaurines." Cretaceous Research 104520 (advance online publication) [[doi: https://doi.org/10.1016/j.cretres.2020.104520]]. | ||
Stromer, E. (1914) "Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. I. Die Topographie und Geologie der Strecke Gharaq-Baharıje nebst Ausführungen über die geologisiche Geschichte-Ägyptens" Abhandlungen der Königlich Bayerischen Akademie der Wissenschaften, Mathematisch-physikalische Klasse, 16(11), 1-78. | Stromer, E. (1914) "Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. I. Die Topographie und Geologie der Strecke Gharaq-Baharıje nebst Ausführungen über die geologisiche Geschichte-Ägyptens" Abhandlungen der Königlich Bayerischen Akademie der Wissenschaften, Mathematisch-physikalische Klasse, 16(11), 1-78. | ||
| Linia 370: | Linia 510: | ||
[[Kategoria:Spinosauridae]] | [[Kategoria:Spinosauridae]] | ||
[[Kategoria:Afryka]] | [[Kategoria:Afryka]] | ||
| − | |||
[[Kategoria:Egipt]] | [[Kategoria:Egipt]] | ||
| − | |||
[[Kategoria:Maroko]] | [[Kategoria:Maroko]] | ||
| − | |||
| − | |||
| − | |||
[[Kategoria:Mezozoik]] | [[Kategoria:Mezozoik]] | ||
[[Kategoria:Kreda]] | [[Kategoria:Kreda]] | ||
[[Kategoria:Cenoman]] | [[Kategoria:Cenoman]] | ||
| − | |||
Aktualna wersja na dzień 20:34, 15 cze 2024
| Autor: | Maciej Ziegler, Kamil Kamiński, Tomasz Sokołowski |
| Korekta: | Tomasz Skawiński, Marcin Szermański |
| Spinosaurus (spinozaur) | |||||
|---|---|---|---|---|---|
| Długość | > 11-13 m / 12-14 m | ||||
| Masa | 3,2-4,2 t / 5,4-7,1 t | ||||
| Miejsce | Egipt (formacja Bahariya) | ||||
| Czas | |||||
| Systematyka | Dinosauria | ||||
 Rekonstrukcja polującego w wodzie spinozaura. Autor: Francesco Delrio [50].
Porównanie wielkości Spinosaurus i człowieka. | |||||
| Mapa znalezisk | |||||
Wczytywanie mapy…
| |||||


Spis treści
Wstęp
Spinosaurus to rodzaj teropoda (spinozauryda), który żył w "środkowej" kredzie na terenach obecnej północnej Afryki. Jedną z cech wyróżniających spinozaura, od której wziął swoją nazwę, były bardzo wysokie wyrostki kolczyste kręgów grzbietowych. Opisanie nowego okazu w 2014 r., a także publikacja dodatkowych informacji w 2020 r. wskazuje, że był to teropod o niezwykłej budowie i biologii oraz pierwszy znany półwodny nieptasi dinozaur (Ibrahim i in., 2014).
Etymologia
Nazwa Spinosaurus oznacza "kolczasty jaszczur" i jest zlepkiem łacińskiego słowa spina, czyli "kolec", oraz greckiego sauros, czyli "jaszczur" (Creisler, online 2003), i odnosi się do wysokich wyrostków kolczystych kręgów tego teropoda (Stromer, 1915). Epitet gatunkowy nawiązuje do Egiptu - kraju, w którym znaleziono jego pierwsze szczątki.
Spinosaurus dał nazwę nowej rodzinie Spinosauridae utworzonej przez Stromera w 1915 roku a później także nadrodzinie Spinosauroidea i podrodzinie Spinosaurinae.
Filogeneza
Zob. Spinosauridae#Filogeneza i budowa.
Historia odkryć


Spinosaurus aegyptiacus to pierwszy rodzaj i gatunek egipskiego dinozaura, odkryty w osadach formacji Baharija 1912 roku i opisany przez niemieckiego paleontologa Ernsta Stromera w 1915 r. Kolejne, dość fragmentaryczne, szczątki z osadów tej formacji, składające się z kręgów i kości kończyn tylnych, Stromer (1934) opisał jako „Spinosaurus B”. Uznał, że mają wystarczająco wiele odmiennych cech, by utworzyć nowy gatunek w obrębie tego rodzaju, lecz nie zdecydował się na to. Obecnie uznaje się, że przynajmniej część z tego materiału należała do innego spinozauryda - Sigilmassasaurus (Evers i in., 2015). Oba okazy zostały zniszczone podczas nalotu bombowego na Monachium w czasie II wojny światowej, w nocy z 24 na 25 kwietnia 1944 r.
W 1996 roku Dale Russell opisał nowy gatunek spinozaura - S. maroccanus - na podstawie kości odkrytych w marokańskich osadach Kem Kem. Za cechę odmienną uznał nieznacznie bardziej wydłużone kręgi szyjne. Jednak w dalszych pracach (Sereno i in., 1998; Rauhut, 2003; Dal Sasso i in., 2005) uznawano drugi gatunek za nomen dubium. Różnice proporcji kręgów Mortimer (online; cytowane przez Hendrickxa i in. [2016]) tłumaczył ich miejscem w kręgosłupie i synonimizował gatunek marokański z egipskim (ale zobacz dalej).
Ważny okaz - MSNM V4047 - przednia część czaszki - ujrzał światło dzienne w 2005 r. Skamieniałość pochodzi z Kem Kem w Maroko (dal Sasso i in., 2005). Jej identyfikacja jako Spinosaurus jest jednak wątpliwa (zob. dalej).
Przełomowe znaczenie miała praca opublikowana w 2014 r. (Ibrahim i in.), w której pobieżnie opisano FSAC-KK 11888 - kolejny okaz z Maroka, obejmujący m.in. kości czaszki, kręgi oraz prawie wszystkie kości miednicy i kończyn tylnych. FSAC-KK 11888 rzucił nowe światło na budowę i tryb życia tego zwierzęcia, ukazując go jako półwodnego drapieżnika o skróconych kończynach tylnych. Publikacja wzbudziła wiele kontrowersji – zob. #Kontrowersje związane z FSAC-KK 11888. Ibrahim i in. (2014) uznali Spinosaurus maroccanus i Sigilmassasaurus brevicollis za synonimy Spinosaurus aegyptiacus. Jednak rok później Evers i in. wykazali, że Sigilmassasaurus brevicollis to ważny gatunek, odmienny od Spinosaurus aegyptiacus, i dokonali jego synonimizacji ze Spinosaurus maroccanus. Obecnie jedynym gatunkiem spinozaura jest więc typowy S. aegyptiacus.
W 2020 r. Ibrahim i in. podtrzymali swoje zdanie na temat synonimizacji S. aegyptiacus z Sigilmassasaurus brevicollis i Spinosaurus maroccanus. Ujawniono też nowe szczątki okazu FSAC-KK 11888, które po raz kolejny zmieniły nasze wyobrażenie o spinozaurze. Okazało się m.in. że ogon był znacznie wyższy niż dotąd sądzono. W tym samym roku Smyth i in. dokonali przeglądu niemal całego materiału kopalnego spinozaurydów z Kem Kem i nie znaleźli podstaw do wyrózniania dwóch taksonów, tym samym kwestionując odrębność sigilmassazaura.
Materiał kopalny

Holotyp
Holotyp to IPHG 1912 VIII 19, opisany przez Stromera w 1915 r. i zniszczony podczas II wojny światowej. Okaz składał się z: fragmentu kości szczękowej, przedniej części żuchwy (kości zębowej, kości płatowej i ?kątowej), 15 izolowanych zębów, łuku neuralnego ?3 (Evers i in., 2015) kręgu szyjnego, 5(?4) (Evers i in., 2015) kręgu szyjnego, czterech (Stromer, 1915; Smith i in., 2006) trzonów kręgów grzbietowych i siedmiu wyrostków kolczystych, kilku żeber i gastraliów, trzech trzonów kręgów krzyżowych i jednego wyrostka kolczystego (Stromer, 1915).
Kontrowersje związane z holotypem
Przypuszcza się, że kręg ogonowy wchodzący w skład holotypu pochodzi od innego taksonu (Stromer, 1934; Rauhut, 2003, s. 35). W 2003 roku Oliver Rauhut zasugerował, że holotyp spinozaura był chimerą allozauroida (kręgi grzbietowe) i teropoda podobnego do Baryonyx (żuchwa i zęby). Nie zostało to jednak przyjęte w późniejszych badaniach (Holtz i in., 2004; Dal Sasso i in., 2005). W 2015 r. Evers i in. (w tym Rauhut) uznali, że skoro Ichthyovenator ma podobnie wydłużone wyrostki kręgów, to przychylają się do konsensusu panującego w tym względzie. Cau (online 2009) wskazał, że kręgi są najbardziej zbliżone do Baryonyx i Megaraptor (uważanego wtedy przez niego za spinozauryda), co potwierdzać miało bliskość filogenetyczną kręgów i żuchwy.


FSAC-KK 11888
FSAC-KK 11888 jest bardziej zbliżony do holotypu Spinosaurus niż do Sigilmassasaurus. Dwa pierwsze okazy mają charakterystyczne poszerzenia (“wybrzuszenia”) w dolnej części kręgów grzbietowych, co różni je od okazu Sigilmassasaurus - „Spinosaurus B” (Stromer, 1934; Evers i in., 2015). Uzasadnia to przypisanie go do Spinosaurus.
FSAC-KK 11888 zawiera fragmenty łba, po kilka kręgów z każdej części kręgosłupa, dwa paliczki dłoni oraz niemal kompletne: obręcz miedniczną i kończyny tylne.

Z publikacji Ibrahima i in. (2014) nie wynika jednoznacznie, które elementy, zwłaszcza kręgosłupa, są obecne. Jeśli nie podano inaczej, cytowanym źródłem jest Ibrahim i in. (2014); skróty odnoszą się do pozycji kręgów: C – szyjny (ang. cervical), D – grzbietowy (ang. dorsal), S – krzyżowy (ang. sacral), Ca – ogonowyn (ang. caudal). Wydaje się, że w skład FSAC-KK 11888 wchodzą niżej wymienione kości. Część kości nosowej, kość przedczołowa, łuskowa, kwadratowo-jarzmowa (niezilustrowana?), większość obu kości kwadratowych, części kości zębowej, izolowane zęby, kość ?łzowa (suplement, s. 2-3). 2-?4 kręgi szyjne (podano wymiary dwóch trzonów, zidentyfikowanych jako C2 i C7 - suplement, s. 32 – tab. S2, lecz ryc. S3 sugeruje, że są trzy trzony - poza ww., też C3 i C9). Min. 5 trzonów kręgów grzbietowych (kręg widoczny na ryc. S2C to D1 wg ryc. S3, choć wg Eversa i in. [2015] kręgów szyjno-grzbietowych nie ma wśród odlewów FSAC-KK 11888 w University of Chicago; D4 i D5 są widoczne na ryc. S3; D5 – ryc. S2E i S3; D6, D7, D8 – wg suplementu s. 32 – tab. S2; wymiary podano tylko dla D6-D8), min. 1 kompletny wyrostek kolczysty (D8 – ryc. 2E) i kilka niekompletnych (Cau, online 2014A; ryc. S3 i ryc. w National Geographic [online 2014] sugerują, że elementy wyrostków zachowane w FSAC-KK 11888 to D4, D5, D6, D7 i D9; równie dobrze mogą one pochodzić z innej części kręgosłupa, np. krzyżowej). Trzony kręgów krzyżowych: S3, S4 i S5 (suplement, tab. 2). ?9 kręgów ogonowych (ryc. S3; nie podano wymiarów żadnego z nich). Żebra szyjne, grzbietowe i min. jedno brzuszne, min. 3 szewrony. Paliczek dłoni II-2 i bliższą część paliczka II-3. Kości miednicy i kończyn tylnych - brak kości pięty, paliczka II-2, kości śródstopia II i IV; nie wiadomo czy jest pazur I.
w 2018 r. zorganiowano kolejną ekspedycję, której owocem było odkrcie kolejnych kości okazu FSAC-KK 11888. Składa się na nie ponad 30 kręgów ogonowych, stanowiących ok. 80% całości ogona. odkryto też dodatkowe elementy czaszki i stopy (Ibrahim i in., 2020b).
Uwaga do ilustracji. Z pokrywających się w holotypie i FSAC-KK 11888 elementów oznaczono te zilustrowane i niefragmentaryczne (np. z FSAC-KK 11888 znane są części kości zębowej). Identyfikacja kości łzowej wg Ibrahima i in. (2014) niepewna. Niezaznaczone są: zęby, żebra szyjne i grzbietowe oraz gastralia (wszystkie są obecne w obu okazach). Nie uwzględniono fragmentów wyrostków kolczystych (wg. ryc. S3 D4-7 i D8). Pominięto również zduplikowane elementy kończyn FSAC-KK 11888. Kości tylnej części czaszki nie zaznaczano osobno. Trzony kręgów szyjnych i grzbietowych i ich wyrostki kolczyste zaznaczono osobno: trzony pod nimi a wyrostki nad nimi. Trzony kręgów krzyżowych oznaczono jednak nad wyrostkami; pierwszy kręg zawiera w holotypie część wyrostka. Kręgi ogonowe, niezależnie od tego, czy zawierają trzony czy wyrostki, oznaczono nad nimi. Identyfikacja elementów FSAC-KK 11888 wg Ibrahim i in., 2014 a holotypu wg Smith i in., 2006 i Stromer, 1915. Ryc. S3 z Ibrahim i in. (2014) jest mało przydatna ze względu na podobieństwo kolorów użytych do rozróżnienia obu okazów (o ile odróżnienie koloru żuchwy czy kości kończyn nie nastręcza trudności, to np. przy kręgach grzbietowych nie jest to proste; ponadto np. kość znajdująca się w tylnej części żuchwy – kątowa, choć jest wyraźnie oznaczona kolorem FSAC-KK 11888, należy do holotypu). Rysunki z National Geographic, online 2014 nie odpowiadają w części ryc. S3 z Ibrahim i in. (2014) (np. tylny kręg szyjny FSAC-KK 11888 zakolorowany jest w całości, podczas gdy wyrostek kolczysty nie jest zakolorowany w ryc. S3; pominięto tylne kręgi szyjne i przednie grzbietowe; D6 – w National Geographic [online 2014] środkowa część wyrostka pochodzi od FSAC-KK 11888, tak jak w ryc. S3, lecz wg tej drugiej górna i dolna część wyrostka pochodzi od holotypu a w rzeczywistości holotyp ma cały wyrostek; zaznaczono tylko dwa kręgi ogonowe FSAC-KK 11888, podczas gdy w ryc. S3 widocznych jest 9, Ibrahim i in. [2014, s. 1613] podają, że FSAC-KK 11888 zawiera tylne kręgi ogonowe; 4-ty kręg ogonowy w holotypie jest kompletny a na ryc. S3 i w National Geographic [online 2014] wydaje się, że trzon jest z FSAC-KK 11888). Kształt wyrostków kolczystych holotypu w znacznej mierze nie pokrywa się ze zdjęciem ze Smith i in., 2006 i rycinami ze Stromer, 1915. Wyrostek kolczysty obecny w holotypie początkowo zidentyfikowano jako C2, lecz Evers i in. (2015) sugerują, że to C3. Ponieważ Ibrahim i in. (2014) podają wymiary trzonu C2, to przyjęto, że również wg nich kręg holotypu to C3 (również w National Geographic [online 2014] jest taka identyfikacja).
Kontrowersje związane z FSAC-KK 11888



Próba ustanowienia FSAC-KK 11888 neotypem Spinosaurus aegyptiacus przez Ibrahima i in. (2014) została skrytykowana przez Eversa i in. (2015). Autorzy ci zwracają uwagę, że znaczna część tego okazu nie została odkryta przez Ibrahima i współpracowników, lecz odkupiona od prywatnego kolekcjonera. Część szkieletu została nabyta przez Uniwersytet w Casablance, a część przez Muzeum Historii Naturalnej w Mediolanie (Cau, online 2014A). Dopiero później zlokalizowano kolekcjonera, który doprowadził autorów opisu do miejsca znalezienia kości, dzięki czemu odkryto dalsze pozostałości (nie wiadomo jednak które). Brak więc danych o powiązaniu kości ze sobą. Evers i in. (2015) zwracają uwagę, że większość kręgowców z osadów Kem Kem jest znajdowanych w wielogatunkowych złożach. Zatem – jeśli nawet zostanie udowodnione, że cały materiał pochodzi z jednego stanowiska, to powinno się następnie wykazać, że są to szczątki jednego osobnika. Jako przesłanki za taką interpretacją autorzy wskazują zbliżoną wielkość i budowę kręgów oraz pasujące do siebie kości udowe i piszczele. Jednak kolekcjonerzy i handlarze skamieniałości z Maroka często kompletują szkielety, zawierające kości z różnych stanowisk, tak, aby do siebie pasowały. Tym samym nie można wykazać, że wszystkie kości wchodzące w skład FSAC-KK 11888 pochodzą od jednego osobnika (Evers i in., 2014). Jak jednak zauważają sami autorzy, występowanie podobnych nietypowych proporcji (znaczne skrócenie kończyn w porównaniu do kręgów) u dwóch okazów (FSAC-KK 11888 oraz u IPHG 1922 X45 - „Spinosaurus B”, prawdopodobnie Sigilmassasaurus) jest zastanawiające. Trudno uznać to za przypadek. Za pochodzeniem kości od jednego okazu świadczyć też mogą cechy młodociane obecne w wielu kościach (Ibrahim i in., 2014). Cau (online 2016A) sugeruje, że za hipotezą, że to jeden osobnik przemawia identyfikacja kości kwadratowej jako pochodzącej od młodego okazu (ang. subadult) przez Hendrickxa i in. (2016). Jednak kość FSAC-KK 11888 jest większa niż inne w tym samym i kolejnym stadium rozwoju, co świadczy przeciwko powiązaniu.
W środowisku paleontologicznym nie ma pełnej zgody co do poprawności wniosków Ibrahima i in. (2014), opartych na budowie FSAC-KK 11888 oraz na będącej podstawą wielu ich hipotez rekonstrukcji, zawierającej szczątki tego oraz innych okazów. Scott Hartman (online 2014A) oraz Mark Witton (online 2014B) wskazywali, że rekonstrukcja FSAC-KK 11888 przedstawia błędnie wyskalowaną miednicę i nogi spinozaura - wg nich są o 27% za małe. Autorzy badania przekonująco wyjaśnili, że rozbieżności wynikają z innych punktów pomiaru w przypadku kręgów a w przypadku kości udowej także z jej niezupełnie pionowego ułożenia (Ibrahim, Maganuco, Keillor i Fabbri w Hartman, online 2014B; Maganuco w Witton, online 2014B).
Ibrahim i in. (2014) zaliczyli do Spinosaurus aegyptiacus szczątki znane jako „Spinosaurus B” (IPHG 1922 X45). Za ich tożsamością mają świadczyć m.in. podobne proporcje długości kręgów do kończyn u IPHG 1922 X45 i FSAC-KK 11888 a także liczne podobieństwa, uwidocznione w ryc. S2 suplementu publikacji Ibrahima i in. (2014). Evers i in. (2015) wskazali jednak na różnice w budowie IPHG 1922 X45 i FSAC-KK 11888, dotyczące m.in. kończyn tylnych, i wykazali, że przynajmniej część tego materiału (kręgi) należy do Sigilmassasaurus (co było jednak później kwestionowane przez Ibrahima i in. [2020] oraz Smytha i in. [2020]). Jeśli więc IPHG 1922 X45 i FSAC-KK 11888 reprezentują ten sam takson a ten pierwszy różni się od holotypu spinozaura (konkretnie kręgów), to powstaje pytanie, czy ten ostatni faktycznie należy do Spinosaurus aegyptiacus (tak też wcześniej Mortimer, online 2014) (art. 75.3.5 ICZN). Jak wyżej wskazano, FSAC-KK 11888 i holotyp spinozaura łączy budowa wyrostków kręgów, jak również mają inne cechy nieprzystające do materiału Sigilmassasaurus (Evers i in., 2015). Autorzy podkreślają znaczną odległość między miejscem znalezienia typowego okazu spinozaura i FSAC-KK 11888 (art. 75.3.6 ICZN), a także wyraźne różnice między Kem Kem (Maroko) i Baharija z (Egipt) (zob. też Carcharodontosaurus#Środowisko). Może się okazać, że FSAC-KK 11888 i inne szczątki spinozaurydów z Kem Kem, odmienne od Sigilmassasaurus, reprezentują nowy, nienazwany jeszcze gatunek. Ponadto Evers i in., jak wcześniej Headden (online 2014), kwestionują konieczność wyznaczania neotypu (art. 75.3 in principio ICZN), gdyż holotyp spinozaura, pomimo iż został zniszczony, to jest stosunkowo dobrze opisany i zilustrowany w literaturze a cechy diagnostyczne są widoczne. Ponadto są dostępne jego zdjęcia (Smith i in., 2006).
Publikacja Ibrahima i in. (2014) budzi wiele innych zastrzeżeń. Jak wskazano wyżej, nie wiadomo, jaki materiał wchodzi w skład okazu (zob. wyżej - #FSAC-KK 11888). Autorzy nie podali, w jaki sposób mierzyli okazy ani jak wykonali rekonstrukcję, która posłużyła np. do wyznaczania środka masy zwierzęcia (np. Witton, online 2014A).


Ibrahim i in. (2020a) bronili swoich wniosków w kolejnej publikacji. Zwrócili uwagę, że FSAC-KK 11888 nie zawiera żadnych podwójnych kości, co powinno mieć miejsce, gdyby szczątki pochodziły od kilku zwierząt. Ponadto parzyste kości z obu stron ciała mają w zasadzie identyczne rozmiary i budowę, a część kości dłoni zachowała się w naturalnym ułożeniu. Ibrahim i współpracownicy uznali również, że stan zachowania kości wskazuje, że przynajmniej część miednicy i kończyn tylnych także była artykułowana w momencie ich znalezienia. Bronili oni także zasadności wyznaczenia okazu FSAC-KK 11888 na neotyp.
Publikacja Ibrahima i in. (2020b) ujawniła dodatkowy materiał okazu FSAC-KK 11888, co w opinii autorów wzmocniło ich konkluzje na temat tego, że szczątki te pochodziły od jednego osobnika. Dzięki odkryciu dodatkowych kręgów ogonowych, możliwe było porównanie z materiałem "Spinosaurus B". Wykazało ono bardzo duże podobieństwo obu okazów pod względem budowy kręgów ogonowych.
Smyth i in. (2020) również postulowali zasadność wyznaczenia neotypu i uznali FSAC-KK 11888 za pewny materiał Spinosaurus aegyptiacus. Kwestionowali m.in. podnoszony przez Eversa i współpracowników argument odległości geograficznej, gdyż ci sami autorzy sugerowali, że część materiału "Spinosaurus B" (Egipt) należy do sigilmassazaura, którego holotyp pochodzi z Maroka.
Materiał przypisany
Badania Eversa i in. (2015), Hendrickxa i in. (2016) oraz Ardena i in. (2018) wskazują, że w cenomanie na obszarze dzisiejszej Afryki Północnej – zarówno w Maroku, jak i w Egipcie, żyły dwa gatunki spinozaurydów – prawdopodobnie Spinosaurus aegyptiacus i Sigilmassasaurus brevicollis. Oba są znane fragmentarycznie, więc trudno oznaczyć większość znalezisk. Chiarenza i Cau (2016) uznali, że jako pewny materiał Spinosaurus aegyptiacus powinno się traktować wyłącznie holotyp, a szczątki spinozaurydów z cenomanu Afryki Północnej, których nie można niewątpliwie przypisać do któregoś z dwóch taksonów, powinny być przypisywane tylko do Spinosaurinae do czasu gruntownej rewizji ich taksonomii.

Prawdopodobnie wszystkie poniższe szczątki pochodzą z osadów Kem Kem z Maroka:
- Cztery kręgi opisane przez Eversa i in. (2015): NHMUK PV R 16429 – przedni kręg szyjny, BSPG 2006 I 57 i BSPG 2013 I 97 – dwa środkowe kręgi szyjne oraz NHMUK PV R 36637 – tylny środkowy kręg szyjny. Autorzy wspomnieli też o okazie ROM 65537 – środkowym kręgu szyjnym. Evers i in. (2015) określali je jako „kręgi drugiego spinozauryda z Kem Kem” a ponieważ różnią się od Sigilmassasaurus, to prawdopodobnie należą do Spinosaurus. BSPG 2006 I 57 ma cechy zbliżające go do FSAC-KK 11888 (Evers i in., 2015).
- Dwa środkowe kręgi szyjne, opisane początkowo jako paratypy S. maroccanus (CMN 41768, CMN 50790). Tak samo jak powyższe, Evers i in. (2015) uznali je za bliżej nieokreślonego spinozauryda. Mortimer zaliczył je do Spinosaurus (tj. jako „aegyptiacus morphotype”). Jako że różnią się one od Sigilmassasaurus (Russell, 1996, ryc. 4d i 7), powinno się je przypisać do Spinosaurus.
- Pięć kości kwadratowych, opisanych przez Hendrickxa i in. (2016), pochodzących od osobników w różnych fazach rozwoju osobniczego (MHNM.KK374, MHNM.KK375, MHNM.KK377, MHNM.KK378 oraz MSNM V6896). Hendrickx i in. (2016) zaliczyli je do spinozaura ze względu na większe podobieństwo do FSAC-KK 11888 niż do drugiego morfotypu kości kwadratowych z Kem Kem.
- Pazur stopy bardzo młodego osobnika (MSNM V6894), mierzącego ok. 1,8 m, który bardzo mocno przypomina pazury FSAC-KK 11888 (Maganuco i Dal Sasso, 2018).
- Dwie kości czołowe, w tym prawa niekompletna (FSAC-KK-3209); dwie kości czołowe i ciemieniowe (FSAC-KK-3210) wstępnie przypisane do Spinosaurus przez Ardena i in. (2018). Reprezentują one mniejszy z dwóch morfotypów wyróżnionych przez autorów. Wg Ibrahima i in. (2020a) nie należą one do spinozaurydów, lecz najprawdopodobniej do krokodylomorfów.
- Kręg szyjny C5 lub C6 (FSAC-KK-7280) oraz przedni kręg grzbietowy (FSAC-KK-18118) przypisane przez Lakin i Longricha (2018). Oba kręgi nie wykazują cech diagnostycznych Sigilmassasaurus, mają za to cechy właściwe dla holotypu spinozaura.
- Sześć kręgów oznaczonych numerami FSAC-KK-5017 do FSAC-KK-5022 (pierwsze pięć to przednie i środkowe kręgi szyjne, ostatni - przedni grzbietowy, prawdopodobnie D2), przypisanych przez Smytha i in. (2020).
Materiał wątpliwy

W przeszłości wiele okazów spinozaurydów zaliczano do Spinosaurus, lecz obecnie wiadomo, że mogą należeć równie dobrze do Sigilmassasaurus. Są to głównie fragmenty czaszki oraz liczne zęby. Najważniejszym z nich jest MSNM V4047 (kości przedszczękowe i niekompletne kości szczękowe i nosowe), opisany przez włoskiego paleontologa Cristiano Dal Sasso z Muzeum Historii Naturalnej w Mediolanie i innych naukowców w 2005 roku. MSNM V4047 jest największym jak dotąd odkrytym osobnikiem spinozauryda (Dal Sasso i in., 2005; Hednrickx i in., 2016) (zob. jednak Sigilmassasaurus#Rozmiary). Sama czaszka mierzyła według Dal Sasso i in. 1,75 metra, a całe zwierzę miało być może około 16-18 metrów długości. W rzeczywistości czaszka mogła być znacznie krótsza - zob. artykuł o Spinosauridae#Wymiary. Ponadto MSNM V4047 ma cofnięte do tyłu nozdrza a prześwietlenie wykazało, że prawdopodobnie pysk mieścił receptory ciśnienia wody (Ibrahim i in., 2014), zob. Spinosauridae#Paleobiologia. Arden i in. (2018) uznali za bardzo prawdopodobne, że okaz ten należał do Sigilmassasaurus, ich zdaniem większego z dwóch spinozaurydów z cenomanu Afryki Północnej. Wg Lacerdy i in. (2021) MSNM V4047 na chwilę obecną nie może być definitywnie przypisywany do Spinosaurus.
Do Spinosaurus może też należeć kość kwadratowa (FSAC-KK-18120) opisana przez Lakin i Longricha (2018). Reprezentuje ona morfotyp, który Hendrickx i in. (2016) zaliczyli do omawianego tu rodzaju. Lakin i Longricha nie zdecydowali się jednak na przypisanie jej do konkretnego spinozauryda z powodu braku materiału czaszkowego w holotypie spinozaura oraz niejasności związanych z FSAC-KK 11888. Podobnie postąpili z kręgiem szyjnym (FSAC-KK-18119) oraz przednim kręgiem piersiowym (FSAC-KK-18121), które wykazują cechy zarówno Spinosaurus jak i Sigilmassasaurus. Ponadto, jako Spinosauridae indet. autorzy ci opisali prawą kość przedszczękową (FSAC-KK-7281), która przypomina okaz MSNM V4047, mogący należeć do sigilmassazaura. Podobnego zdania są Lacerda i in. (2021).
Spinozaurowi przypisywano też szczątki z cenomańskiej formacji Echkar w Nigrze. Należy to nich kręg grzbietowy, początkowo przypisany do Carcharodontosaurus iguidensis, fragmentaryczny pysk (MNBH EGA1) oraz pojedyncze zęby. Sereno i in. (2022) zaznaczają, że szczątki te przypominają materiał S. aegyptiacus, lecz mogą należeć do innego gatunku. Równie prawdopodobne wydaje się, że może to być odrębny rodzaj spinozauryda.
Najnowsze publikacje (Ibrahim i in., 2020a; Ibrahim i in., 2020b; Smyth i in., 2020) wskazują na synonimikę Sigilmassasaurus i Spinosaurus, a różnice między nimi uznaje się w nich za wynik zmienności osobniczej, możliwego dymorfizmu płciowego, ontogenezy, pośmiertnych uszkodzeń szczątków, różnego stanu ich zachowania oraz porównywania ze sobą różnych kości, zwłaszcza kręgów, które mogły cechować się zmienną morfologią w zależności od ich położenia w kręgosłupie, nawet u pojedynczego osobnika. Tym samym cały materiał przypisywany do sigilmassazaura należałby do Spinosaurus. Na obecną chwilę wydaje się, że kwestia ta wymaga jeszcze dalszych badań.
Budowa i paleobiologia
Plan ciała, budowa i paleobiologia ogólnie

Czaszka Spinosaurus była prawdopodobnie wydłużona, jak u innych spinozaurydów (zob. niżej - #Czaszka). W przeciwieństwie do nich, długi był też tors, na którym znajdował się wysoki grzebień (zob. niżej - #Żagiel). O ogonie niewiele można stwierdzić – jak u innych teropodów, był zapewne długi i silnie umięśniony. Opierając się na krewniakach, uznać należy, że kończyny przednie były zapewne dobrze rozwinięte i pomocne w łapaniu ryb. Ibrahim i in. (2014) wspomnieli, że dłonie były proporcjonalnie większe niż u wcześniejszych spinozaurydów, nie podali jednak dokładniejszych danych. Kończyny tylne były zredukowane i słabo przystosowane do dźwigania ciężaru ciała (zob. niżej - #Kończyny tylne).
Ibrahim i współpracownicy (2014) dowiedli, że co najmniej kości udowe i wyrostki żagla FSAC-KK 11888 były zbudowane głównie z gęstej tkanki kostnej, ze zmniejszoną jamą szpikową. Gęstość kości była 30-40% większa niż u innych teropodów i podobna jak u pingwinów królewskich. Interpretuje się to jako przystosowanie do półwodnego trybu życia (Ibrahim i in., 2014).
Spinosaurus był niezwykłym zwierzęciem, diametralnie różnym nie tylko od innych teropodów, ale także pozostałych spinozaurydów. Dotyczy to zwłaszcza nietypowych proporcji kończyn tylnych i jego domniemanej czworonożności, czym różnił się choćby od Suchomimus. Mimo pewnych zastrzeżeń (zob. #Kontrowersje związane z FSAC-KK 11888), trudno zaprzeczyć, że Spinosaurus był dobrze przystosowany do wodnego trybu życia i rybożerności.
W tyn kontekście zaskakujące są wyniki badań składu chemicznego zębów. Wynika z nich, że przedstawiciele Spinosauridae spędzali w wodzie znaczną ilość czasu, podobnie jak krokodyle czy hipopotamy. Jednak spinozaurydy z cenomanu Afryki Północnej – w tym na pewno Spinosaurus – okazały się bardziej lądowe niż ich krewniacy (Amiot i in., 2010). Zob. też Spinosauridae#Paleobiologia.
Diagnoza


Ibrahim i in. (2014) podali 10 pozaczaszkowych cech diagnostycznych spinozaura (literą „H” i „F” oznaczono cechy obecne odpowiednio w holotypie i u FSAC-KK 1188, za odpowiednio Stromerem [1915] i Ibrahimem i in. [2014]).
(1) Silnie zwężone i wydłużone trzony kręgów grzbietowych o kształcie klepsydry (H, F [Cau, online 2014A]) – powodowało to znaczne wydłużenie grzbietu zwierzęcia.
Kolejne cechy odnoszą się do tworzących „żagiel” wyrostków kolczystych kręgów grzbietowych, które: (2) były do 10 razy wyższe niż wysokość trzonów (H, F); (3) miały największą przednio-tylną szerokość poniżej swych wierzchołków (ang. „below spine apex”) (sic!) – być może chodzi o unikalne rozszerzenia (wypukłości) widoczne w dolnej części wyrostków (H, F); (4) były zbudowane z gęstej tkanki kostnej z wąskim pasem tkanki gąbczastej (H, F); (5) były pokryte pionowymi prążkami – w diagnozie (suplement, s. 9) podano, że znajdują się one na bliższej (dolnej) jedną trzeciej wyrostków, lecz w publikacji (s. 1616 i ryc. 4D) wskazano, że są one na dystalnej (górnej) powierzchni; w niektórych kręgach holotypu widoczne jest to na całej wysokości (H, F).
Diagnostyczna jest także budowa kończyn tylnych: (6) kości długie - tj. co najmniej udowa - nie miały otwartej jamy szpikowej (F); (7) długość kości biodrowej była większa niż długość kości udowej (F); (8) kość udowa była silnie wygięta ku przodowi (ang. „bowed anteriorly”), z rozrośniętym czwartym krętarzem, rozciągającym się wzdłuż 25% trzonu tej kości (F); (9) pierwszy palec stopy był długi; paliczek I-1 był najdłuższy z (niepazurowych) paliczków stopy (F); (10) pazury stóp były szersze niż ich głębokość (wysokość), o długości prawie cztery razy większej od bliższej głębokości; pazury miały płaską dolną powierzchnię (F).
Cechy te związane były prawdopodobnie w znacznej mierze z lokomocją (2?, 3?, 7, 8, 9, 10) i z wodnym trybem życia (1?, 3?, 4, 6, 7, 8, 10). Ibrahim i in. (2014) wymienili także dwie cechy pyska, jednak nie odnoszą się one do niewątpliwego materiału Spinosaurus.
Kilka „potencjalnie autapomorficznych cech” obecnych w holotypie wymienili także Evers i in. (2015, s. 74). Spinosaurus z pewnością jest odróżnialny od sigilmassazaura – różni się od niego prawdopodobnie budową kręgów i kończyn (Evers i in., 2015) oraz kości kwadratowej (Hendrickx i in., 2016) (zob. też Sigilmassasaurus#Budowa i paleobiologia). Odmienna była też budowa kości czołowych – te przypisane spinozaurowi były proporcjonalnie dłuższe i węższe (Arden i in. 2018). Ibrahim i in. (2020a) uznali jednak, że kości te nie należały w ogóle do spinozaurydów. Z kolei Smyth i in. (2020) uznali różnice w budowie kręgów i kości kwadratowej za nieistotne taksonomicznie.
Czaszka

Czaszka spinozaura jest znana jedynie z kilku elementów. W zasadzie jedyny pewny a niefragmentaryczny materiał to część żuchwy. Prawdopodobnie miał on długą i niską czaszkę, z przesuniętymi do tyłu nozdrzami i stożkowatymi zębami. Czaszka niezidentyfikowanego spinozauryda MSNM V4047 ma na pysku liczne otwory i kanały – prawdopodobnie mieszczące receptory ciśnienia wody, zob. Spinosauridae#Paleobiologia. Wg Ardena i in. (2018) nietypowa budowa kości czołowych wskazuje na wysokie umiejscowienie oczodołów, ponad resztę czaszki (z czym jednak nie zgodzili się Hone i Holtz [2021]) oraz zredukowane opuszki węchowe. Te ostatnie prawdopodobnie były mało przydatne dla zwierzęcia polującego głównie w wodzie.
Niekompletna żuchwa została opisana przez Stromera (1915). Grzbietowa (górna) krawędź tworzy łuk, wypukły do szóstego zębodołu, dalej ku tyłowi wklęsły do dwunastego zębodołu, który następnie znów staje się wypukły i lekko wznosi się do góry. Brzuszna (dolna) krawędź żuchwy ma kształt zwróconego przeciwnie łuku, który najwyższą wysokość osiąga w okolicach siedemnastego zębodołu. Końcówka żuchwy jest kanciasta i stosunkowo wysoka, nadając jej specyficzny kształt. Pomiędzy trzecim a czwartym zębodołem żuchwa ma 13,5 cm wysokości, w okolicach siódmego – zaledwie 9 cm, na wysokości piętnastego – 15,5 cm, a 10 cm dalej ku tyłowi – aż 19 cm. Jej budowa wskazuje, że Spinosaurus miał długi, wąski pysk. Wg Stromera, w tylnej części kości żuchwy wydają się „cienkie i słabe”. Co ciekawe, obie połówki dolnej szczęki różnią się ilością zębów – prawa ma 15 zębodołów, a lewa – 16. Zęby mocno różniły się wielkością w zależności od położenia w żuchwie. Zęby od drugiego do czwartego, a więc leżące w podniesionej przedniej części żuchwy były wyraźnie większe od pozostałych, po nich następuje seria mniejszych zębów aż do dziesiątego, kolejne cztery znowu stają się większe, a ostatni jest ponownie mniejszy i silniej bocznie spłaszczony. Zęby spinozaura były tylko nieznacznie zakrzywione, a w przekroju poprzecznym – prawie okrągłe. Ich korzenie były niezwykle długie i w górnej części grubsze od koron.
W 2023 r. Smith i Martill opisali niezwykły patologicznie zmieniony ząb z osadów Kem Kem, mogący należeć do spinozaura. Wykazuje on m.in. zwiększoną krzywiznę korony i obecność trzech głębokich rowków wzdluż całej korony.
Kończyny tylne

Budowa kończyn tylnych wg Ibrahima i in. (2014) wskazuje na przystosowanie do półwodnego trybu życia. Były one skrócone a miednica była zredukowana. Jak jednak sugeruje Cau (online 2016C), nie tyle kończyny były pomniejszone, co wydłużony był grzbiet.
W przeciwieństwie do innych dużych teropodów, kość udowa była krótsza od piszczelowej. Stosunkowo długie było także śródstopie. Pod względem proporcji długości kończyn Spinosaurus wydaje się najlepiej przystosowany do szybkiej lokomocji wśród bazalnych tetanurów. Współczynnik obliczony metodą Personsa i Curriego (2016) na podstawie wymiarów podanych przez Ibrahima i in. (2014) to +8%. Wśród niecelurozaurowych teropodów jedynie niektóre ceratozaury go przewyższają (zob. Ceratosauria#Lokomocja). U teropodów proporcje te zmieniają się znacznie w trakcie wzrostu – u młodych kończyny tylne, zwłaszcza dolne części, są proporcjonalnie dłuższe (np. Persons i Currie, 2016); u spinozaura mogło być podobnie. Takie proporcje u Spinosaurusz pewnością nie znamionują jednak szybkiego ruchu, biorąc pod uwagę jak niewielkie są tylne kończyny w porównaniu do szkieletu osiowego (Ibrahim i in., 2014; jak wskazano wyżej nie ma jednak pewności co do pochodzenia szczątków od jednego osobnika). Kość udowa miała masywne miejsca przyczepu mięśni odpowiedzialnych za zginanie kończyny do tyłu. Dalsze kłykcie kości udowej są wąskie a grzebień goleniowy piszczeli średnio rozwinięty. Taka budowa przypomina stan obserwowany u wczesnych waleni i współczesnych półwodnych ssaków, używających łap do wiosłowania. Wydłużenie pierwszego palca stopy (zredukowanego u innych teropodów) i niskie, spłaszczone pazury (podobne do pazurów ptactwa brzegowego) mogły być przystosowaniem do poruszania się po grząskim gruncie i wiosłowania w wodzie. Być może między palcami znajdowała się błona pławna. Co ciekawe, budowa i proporcje pazurów pozostawały prawdopodobnie takie same przez całe życie, ułatwiając pokonywanie miękkich podłoży lub wiosłowanie. Wskazuje to, że tryb życia młodych spinozaurów nie odbiegał zbytnio od dorosłych (Maganuco i Dal Sasso, 2018). Potwierdzają to Lakin i Longrich (2018) którzy sugerują jednak, że bardzo młode osobniki (poniżej 2 m długości) zajmowały nieco inne nisze ekologiczne i występowały w innym środowisku.
Proporcje ciała spinozaura – wydłużony grzbiet i krótkie kończyny – są częste u zwierząt wodnych (Nash, online 2014B). Zbliżoną budowę miał prawdopodobnie bliski krewny spinozaura - Sigilmassasaurus (Evers i in., 2015).
Żagiel
Budowa żagla

Tzw. "żagiel" spinozaura był niezwykły, choć inne dinozaury, takie jak ornitopod Ouranosaurus czy hadrozauryd Barsboldia, wytworzyły podobne struktury kostne. Podobne twory miały także permskie synapsydy, takie jak Dimetrodon i Edaphosaurus. Żagiel jest cechą wytworzoną niezależnie u każdego spośród wymienionych gadów - czyli analogiczną. Wyrostki spinozaura były też najdłuższe wśród dinozaurów – miały do ok. 165 cm wysokości i były do 8 (Stomer, 1915) lub 10 (Ibrahim i in., 2014) razy wyższe od trzonów.
Kształt ozdoby jest niepewny. Tradycyjnie uznawano, że żagiel rozciągał się półkoliście nad grzbietem (Smith i in., 2006). Proponowano również, że najdłuższy wyrostek kolczysty znajdował się na ogonie (Cau, online 2008; Headden, online 2010 [wątpliwości metodologiczne budzą przesłanki 6 i 7 Headdena]). W rekonstrukcji Ibrahima i in. (2014), opartej również na FSAC-KK 11888, żagiel ma trapezoidalny kształt z dwoma wierzchołkami. Wysokość wyrostków kręgów krzyżowych i ogonowych jest w niej spekulatywna. Opisanie w 2020 r. niemal kompletnego ogona wykazało, że żagiel ciągnął się przez cały ogon, przyjmując formę podobną do tworów krokodyli bądź traszek. Wyrostki kolczyste przednich kręgów ogonowych były trzykrotnie wyższe od trzonów kręgów, a tylnych - aż siedmiokrotnie. Wydłużone były również szewrony, co zwiększało boczną powierzchnię ogona. Jednocześnie zredukowane były przed- i zazygapofyzy, co zwiększało giętkość ogona (Ibrahim i in., 2020b).
Stromer (1915) zauważył, że gładka powierzchnia i słaba konstrukcja górnych końców wyrostków przeczy przypuszczeniu, by żagiel był pokryty silnymi mięśniami. Zaznaczył, że taki pogląd nie pasuje też do lokalizacji tej niezwykłej struktury. Największe wyrostki kolczyste znajdowały się jego zdaniem w okolicy lędźwiowej, gdzie nie ma powodu do nadzwyczajnej rozbudowy mięśni grzbietu. Wg Stromera był to skórny żagiel, podobny do tego, który występuje u kameleona grzebieniastego (Chamaeleo cristatus). Obecne u obu gatunków pionowe prążki, ostre brzegi i duże odległości między wyrostkami również zdaniem Ibrahima i in. (2014) sugerują, że struktura spinozaura była pokryta warstwą skóry.
Jack Bailey (1997) uznał, że żagiel nie był delikatną ozdobą pokrytą cienką warstwą skóry, a przedłużeniem grzbietu zwierzęcia, pokrytym mięśniami i ścięgnami. Jego zdaniem budowa wyrostków kolczystych kręgów spinozaura bardziej zbliża go do ssaków mających garb na grzbiecie, jak np. bizon, niż do bazalnych synapsydów, jak Dimetrodon. U dinozaurów i ssaków wyrostki są masywniejsze i w widoku z góry wydłużone, a u bazalnych synapsydów są delikatne i okrągłe w przekroju. Wskazuje się jednak, że taka budowa u spinozaura jest po prostu dziedzictwem po przodkach (Holtz i in., 2004).
Funkcje żagla
Na temat funkcji tzw. "żagla" spinozaura powstało wiele hipotez, przy czym nowsze (od 2014 r.) często wiążą go z półwodnym trybem życia.
Żagiel mógł służyć do przyciągania samic i odstraszania skromniej obdarzonych rywali. Podobnie jak współczesne zwierzęta stosują swoje rogi, poroże lub kontrastową barwę podczas okresu godowego, również i spinozaur mógł używać swojego żagla by zainteresować wybredne samice. Samiec o większym i bardziej okazałym żaglu mógł wydawać się silniejszy lub zdrowszy. Możliwe także, że żagle w czasie zalotów napełniały się krwią w celu lepszej prezentacji.
Inną możliwą funkcją żagla było powodowanie, że zwierzę wydawało się jeszcze większe, niż w rzeczywistości, co mogło mieć znaczenie w konkurowaniu z innymi mięsożercami.

Zdaniem Baileya (1997), struktura ta mogła służyć jako magazyn lipidów i substancji energetycznych (zwłaszcza przy dalekich wędrówkach lub w porze niedoboru pokarmu), pomagać w utrzymaniu gigantotermii lub stanowić osłonę przed gorącem. W związku z dużą masą takiej struktury, Bailey postulował czworonożność spinozaura.

Cau (online 2016C), zaproponował, że u dwunożnego Spinosaurus wysokie wyrostki kolczyste kręgów były przystosowaniem do wydłużonego grzbietu. Cau posłużył się przykładem jednoprzęsłowego mostu budowanego z jednego brzegu kanionu, na którym umieszczony jest jedyny punkt podparcia. Wyjaśnił, że wraz z dokładaniem kolejnych przęseł (wydłużaniem mostu), podtrzymujące go liny muszą być przyczepione do coraz wyższego pylonu (wieży). W przypadku teropodów „linami” są mięśnie i ścięgna nadosiowe (epaxial) a pylonem wyrostki kolczyste. Wcześniej Cau (online 2013D) sugerował podobną funkcję na przykładzie koparek wielonaczyniowych. Cau (online 2014B), posługując się kręgami szyjnymi Sigilmassasaurus, stwierdził, że Spinosaurus miał pionową, wysoko uniesioną szyję, niemal stykającą się z żaglem. Cau zasugerował, że ścięgna utrzymujące szyję w takim położeniu, bez użycia mięśni, były przyczepione do wyrostków kolczystych kręgów grzbietowych. Jest to problematyczna rekonstrukcja nie tylko z powodu użycia kręgów innego spinozauryda (Spinosaurus mógł być przecież zbudowany podobnie), lecz także ze względu na podważenie rekonstrukcji „U-kształtnej” szyi Sigilmassasaurus Russella (1996) przez McFeetersa i in. (2013). Mortimer w komentarzach do postu Cau nie zgodził się z taką interpretacją a sam Cau niedostatecznie wyjaśnił swój pomysł.
Sugerowano, że żagiel mógł pełnić funkcję termoregulacyjną. Mógł być obficie unaczyniony i używany do nagrzania się w chłodniejsze, lecz nie zachmurzone dni i do ochładzania organizmu w czasie upałów. Mogło mieć to szczególne znaczenie przy ziemnowodnym trybie życia. Gdy dinozaur pływał w wodzie, żagiel zdaniem Ibrahima i in. (2014) wystawał nad jej powierzchnię. Wychwytywał zatem zapewne promienie słoneczne i ogrzewał zwierzę. Jeśli dinozaury, w tym Spinosaurus, miały metabolizm pośredni między gadzim a ssaczym (Grady i in., 2014), to tego typu struktura mogła być niezwykle pomocna w środowisku wodnym, które szybko wychładza organizm. Wcześniej sugerowano, że hipoteza mezotermiczności wyjaśnia, dlaczego wśród dinozaurów nie było zwierząt wodnych (Sampson, 2009). Z pewnością również wielkie rozmiary pomagały spinozaurowi utrzymać ciepłotę ciała w wodzie.
Ibrahim i in. (2014) sugerują, że funkcją pełnej w środku ozdoby było również dociążenie zwierzęcia.
Nash (online 2016) wskazuje, że żagiel mógł pomagać w łowieniu ryb przez zwiększanie powierzchni, którą spinozaur mógł blokować odpływ, uniemożliwiając ucieczkę ofiarom i je stłaczając. Żagiel mógł też służyć do zacieniania powierzchni wody, ułatwiając wypatrzenie zdobyczy.

Inną hipotezę, związaną z podwodną lokomocją, przedstawili Gimsa i in. (2015). Ich zdaniem żagiel był strukturą analogiczną do płetwy grzbietowej żaglicy, której kształt odpowiada zrekonstruowanemu przez Ibrahima i in. (2014). Autorzy wykluczają wyłącznie pokazową funkcję żagla, gdyż byłby on przeszkodą na lądzie, a w wodzie musiałby przetrwać zanurzenie i zapewnić odpowiednią stabilność - w przeciwnym wypadku naraziłby zwierzę na uraz kręgosłupa. Ich zdaniem żagiel był używany przede wszystkim pod wodą i miał głównie hydrodynamiczne funkcje. Gimsa i in. sugerują, że żagiel był stabilizatorem – zapobiegał odchylaniu się tułowia podczas gwałtowniejszych ruchów głowy i szyi, co miałoby pozwalać na większy zasięg i szybkość ruchu łba w pogoni za zdobyczą (autorzy nie pokusili się jednak na dokładniejsze opisanie tego mechanizmu). Ponadto miałby działać podobnie jak miecz w łodzi, tj. zmniejszać znoszenie ciała na boki w wyniku bocznych ruchów ogona. Nash (online 2016) sugeruje, że żagiel pomagał zachowaniu równowagi (zapobiegał zataczaniu się) w skręcaniu i obracaniu się pod wodą. Hipoteza Gimsy i in. (2015) wydaje się prawdopodobna, choć wymaga dalszych badań. Pewne kwestie są jednak problematyczne - kształt żagla nie jest pewny a szyja została zrekonstruowana jako zbyt długa.
Postawa i lokomocja

Poruszanie się na lądzie
Opisanie FSAC-KK 11888 ożywiło pogląd o prawdopodobnej czworonożnej postawie Spinosaurus. Wcześniejsze hipotezy o czworonożności należy uznać za bezpodstawne, ponieważ nie były znane kości kończyn.
Typowa dla teropodów dwunożna lokomocja była w przypadku Spinosaurus co najmniej znacznie utrudniona. Kończyny tylne były zmniejszone. Powierzchnia pełniącej funkcję przyczepu mięśni kości biodrowej była około dwukrotnie mniejsza niż u innych teropodów. Grzebień nadpanewkowy, będący podporą kończyny, był niski. Dźwigająca ciężar ciała artykulacja stawu kolanowego była zredukowana (Ibrahim i in., 2014). Ponadto według rekonstrukcji z ww. publikacji, środek ciężkości był przesunięty do przodu. Zdaniem Ibrahima i in. (2014), uniemożliwiało to dwunożny chód, mogło jednak pomagać w utrzymaniu równowagi podczas pływania.

Wg Cau (online 2015) wysokie wyrostki kolczyste, smukły tors i budowa ogona sugerująca, że Spinosaurus mógł się nim podpierać, wskazują, że dinozaur mógł przyjmować półwyprostowaną, dwunożną postawę. W rekonstrukcji Cau użył również kręgów szyjnych Sigilmassasaurus, które powodowały, że szyja była znacznie odgięta ku tyłowi, dodatkowo przesuwając środek ciężkości w tym kierunku (Cau, online 2014B). Wg Nasha (online 2016) młode okazy mogły biegać na dwóch nogach w półwyprostowanej pozycji.
Pełna czworonożność jest jednak mało prawdopodobna. Jedyny zilustrowany element kończyny przedniej znany u FSAC-KK 11888 (paliczek, kolejny element to część kolejnego paliczka) nie ma budowy sugerującej, że pełnił funkcje podporowe (Ibrahim i in., 2014, ryc. 2H).
Nash (online 2014A) zasugerował, że na lądzie Spinosaurus poruszał się prawdopodobnie przez ślizganie się na brzuchu.
Wg Hone i Holtza (2021) układ wyrostków bocznych ogona wskazuje na ułożenie mięśni kończyn tylnych typowe dla zwierząt lądowych. Nietypowa budowa stopy z szerokim rozstawem palców i użycie piątego palca w celach podporowych może świadczyć o przystosowaniu do poruszania się po lądzie, zwłaszcza po miękkim gruncie, i to pomimo zmniejszonych kończyn. Autorzy ci sugerują, że lokomocja lądowa spinozaura mogła się ograniczać do przemieszczania się między żerowiskami.
Wg Sereno i in. (2022) Spinosaurus na lądzie był zwierzęciem dwunożnym ze środkiem ciężkości umieszczonym nad stopami. Było to możliwe m.in. dzięki znacznej pneumatyzacji szkieletu osiowego w części przedkrzyżowej. Z kolej żagiel grzbietowy i ogon służyły głównie jako twory na pokaz (Sereno i in., 2022).
Poruszanie się w wodzie
Ibrahim i in. (2014) podali, że trzony kręgów ogonowych Spinosaurus były krótkie, a połączenia łuków neuralnych zredukowane, ułatwiając boczne zginanie ogona i wykorzystywanie go jako napędu. Dodatkowym napędem miałyby być stopy. Niepublikowane badania (Sereno i in., 2015) sugerują, że Spinosaurus był wolnym pływakiem.

Nash (online 2016) jako prawdopodobny sposób poruszania się spinozaura w wodzie wskazał nie pływanie, lecz chodzenie podwodne (dobijanie się od dna) na modłę hipopotamów. Wskazywać na to mają gęste kości, zmniejszona miednica i rozrost mięśnia caudofemoralis.
Poruszanie się w wodzie zapewne ułatwiała też specyficzna budowa kości czołowych, przez co oczy spinozaura były położone wyżej niż u innych teropodów i podniesione ponad resztę czaszki. Dzięki temu zwierzę mogło prawdopodobnie obserwować powierzchnię wody pomimo zanurzenia niemal całej czaszki (Arden i in. 2018). Z taką interpretacją nie zgodzili się Hone i Holtz (2021) którzy wykazali, że zachowanie możliwości obserwacji powierzchni wody wymagało wynurzenia niemal całej górnej części czaszki. Podobnie oddychanie nie było prawdopodobnie możliwe bez wystawiania niemal całej głowy, bowiem nozdrza spinozaura były wprawdzie cofniete do tyłu, ale nie skierowane do góry, jak u współczesnych krokodyli czy hipopotamów. Spinzaur bardziej przypominał w tym względzie ptaki brodzące, jak czaple.
W 2018 r. ukazała się praca Donalda Hendersona, podająca w wątpliwość półwodny tryb życia sinozaura. Autor posłużył się trójwymiarowym, cyfrowym modelem zwierzęcia, opartym w głównej mierze na rekonstrukcji z pracy Ibrahima i in. (2014). Został on porównany z modelem współczesnego aligatora oraz kilku innych teropodów, m.in. zuchomima i tyranozaura. Badania wykazały, że środek ciężkości znajdował się znacznie bliżej bioder, niż sądzili Ibrahim i współpracownicy. Dzięki temu spinozaur wciąż byłby w pełni zdolny do poruszania się po lądzie. Autor stwierdził, że Spinosaurus z pewnością był w stanie unosić się na wodzie i oddychać z głową wystawioną nad jej powierzchnię, nie ma w tym jednak niczego specjalnego, gdyż większość zwierząt lądowych jest do tego przystosowana. Henderson wywnioskował, że spinozaur prawdopodobnie nie byłby w stanie całkowicie się zanurzyć z powodu zbyt małej średniej gęstości ciała, nawet po opróżnieniu płuc z powietrza w 75%. Stanowiłoby to bardzo dużą przeszkodę w ściganiu i chwytaniu wodnych zwierząt. Model skonstruowany przez Hendersona pokazał również, że utrzymanie pionowej postawy w wodzie byłoby bardzo trudne bez opierania się na kończynach, gdyż ciało spinozaura miałoby tendencję do przechylania się na bok. Wreszcie, podkreślił on niepewność co do poprawności rekonstrukcji w pracy Ibrahima i współpracowników. Wszystko to sprawia, że autor uznał spinozaura za teropoda wprawdzie mocno związanego z wodą, ale jego zdaniem nie było to wysoce wyspecjalizowane zwierzę, prowadzące półwodny tryb życia, ale raczej brodzące w płyciznach i aktywne również na lądzie. Do podobnych wniosków doszli Hone i Holtz (2021). Zauważyli oni, że silne wygięcie w dół szyi spinozaura, kształt czaszki, umiejscowienie nozdrzy i oczodołów predysponują go do polowania na linii brzegowej, z pyskiem częściowo zanurzonym lub znajdującym się tuż nad powierzchnią. Dzięki temu był on w stanie chwytać przepływające ryby za pomocą szybkich, pionowych ruchów głowy. Budowa zębów spinozaura wskazuje, że żywił się stosunkowo twardą wodną zdobyczą, taką jak ryby pancerne, skorupiaki i żółwie.
Ibrahim i in. (2020b) na podstawie opisu nowych szczątków podtrzymali swoje zdanie o spinozaurze jako zwierzęciu spędzającym większość czasu w wodzie. Głównym napędem byłby ogon, zbudowany inaczej niż u reszty teropodów. Dzięki zwiększonej powierzchni bocznej i dużej elastyczności, byłby on znacznie bardziej efektywnym napędem w wodzie niż ogony celofyza i allozaura, z którymi go porównywano. Ustępował jednak w tym względzie ogonowi współczesnego krokodyla nilowego.
Hone i Holtz (2021) wykazali, że ogon spinozaura był stosunkowo sztywny u nasady a elastyczny na dalszym odcinku, co zdaje się podważać jego funkcje lokomotoryczne i wskazuje bardziej na funkcje związane z doborem płciowym. Autorzy ci uznali, że omawiany teropod był w stanie pływać, zapewne nawet lepiej niż inne teropody, ale raczej nie był zwierzęciem aktywnie polującym pod wodą. Takie zachowania byłyby także mocno utrudnione przez grzbietowy żagiel i stosunkowo duże kończyny przednie. Również badania Fisha i in. (2021) wykazały, że ogon spinozaura wytwarzał w wodzie znacznie mniejszy ciąg niż u współczesnego krokodyla.
Wg Beevora i współpracowników o wodnym trybie życia spinozaura świadczy też powszechne występowanie jego zębów w osadach rzecznych. Udział zębów typowo lądowych dinozaurów nie przekracza tam kilku procent, natomiast zęby spinozaura w niektórych miejscach są nawet liczniejsze niż zęby ryby Onchopristis. Hone i Holtz (2021) uznali, że świadczy to o polowaniu spinozaura w rzekach (pod wodą lub na brzegu), ale nie przesądza o tym, czy spędzał on większość czasu zanurzony w wodzie.
Fabbri i in. (2022a) zaobserwowali u spinozaurydów wyraźny wzrost gęstości kości i m.in. na tej podstawie uznali Spinosaurus za nurkującego drapieżnika, zdolnego do polowania pod wodą. Wnioski z tej pracy były podważane przez Myhrvolda i in. (2022), którzy uznali dobór próbek za zbyt selektywny. Wskazywali też na nieścisłości dotyczące struktury kości spinozaurydów i błędne wnioski statystyczne. Ich zdaniem stosunkowo duża gęstość badanych próbek kości może wynikać po prostu ze znacznych rozmiarów ciała spinozaura, a nie przystosowań do nurkowania. Fabbri i in. (2022b) podtrzymali jednak swoje stanowisko w kolejnej publikacji. Myhrvold i in. (2024) ponownie skrytykowali wnioski Fabbriego i współpracowników. Podważyli m.in. możliwość wyciągania daleko idących wniosków z nielicznych i niepewnych szczątków. Ponadto zwrócili uwagę, że ze względu na wysoka pneumatyzację spinozaur prawdopodobnie nie był w stanie nurkować. Ich zdaniem Spinosaurus był półwodnym rybożercą, ale nie drapieżnikiem polującym aktywnie pod wodą.
Do podobnych wniosków doszli Sereno i in. (2022). Wg tych autorów, spinozaur był półwodnym drapieżnikiem polującym w strefie przybrzeżnej, ale nie zwierzęciem ściśle wodnym. Wykonany przez nich model cyfrowy ujawnił, że w głębokiej wodzie Spinosaurus byłby powolnym i niestabilnym pływakiem, niezdolnym do nurkowania z powodu zbyt dużej wyporności. Gęstość jego ciała oszacowano na 830 kg/m3, podczas gdy gęstość słonej wody wynosi 1026 kg/m3.
Wg Smarta i Sakamoto (2024) umiejscowienie nozdrzy spinozaura nie byłoby zbyt pomocne przy całkowicie wodnym trybie życia, jest natomiast użyteczne dla zwierząt takich jak rybożerne ptaki, brodzące w wodzie i polujące na przepływające ryby.
Wymiary

Już od odkrycia pierwszych kości w 1912 roku, Spinosaurus był jednym z najbardziej prawdopodobnych kandydatów do tytułu najdłuższego i najcięższego teropoda. Wszelkie oszacowania sprzed opisania FSAC-KK 11888 w 2014 r. należy jednak pominąć, jako opartych na nieprawidłowych rekonstrukcjach.
Wg Ibrahima i in. (2014) maksymalne rozmiary dorosłego osobnika to 15 m długości. Oszacowanie masy według ich rekonstrukcji wykazało wartość 7.557 kg (Franoys, online 2016); przy identycznych założeniach jak Hartmana (online 2013), który uzyskał dla Tyrannosaurus FMNH PR2081 8.400 kg i dla Giganotosaurus MUCPv-95 8.200 kg. Zwiększenie gęstości kończyn i żagla o hipotetyczne i zapewne zawyżone 20% (por. np. Wedel, 2007; Henderson, 2004) powoduje, że masa zwiększa się o ok. 154 kg - do 7.713 t.
Mortimer (online 2016) podaje, że holotyp mierzył 13,2 m przy długości czaszki 1,45 m. Wg Cau (online 2014C) okaz mierzył 12-13 m a czaszka mogła mieć ok. 1,32 m (Cau, online 2013C; okaz MSNM V4047 Cau szacuje na ok. 1,3 m i czasem na 1,4 m i uważa również, że jest on 5-10% większy od holotypu – zob. Spinosauridae#Wymiary). Cau twierdził także (online 2013A), że okaz jest porównywalnych rozmiarów z holotypem Baryonyx. Wg rekonstrukcji obok długość ciała to ok. 11 m a czaszki - 1,15-1,25 m. Zdania na temat dojrzałości okazu są podzielone. Niektórzy podają (np. Paul, 1988), że był to okaz niedorosły ze względu na niezrośnięte trzony kręgów grzbietowych i ich wyrostki. Inni twierdzą (np. Cau, online 2013C), że okaz jest dorosły. Stromer (1915) wskazał, że w kręgu szyjnym trzon jest „połączony z wyrostkiem neuralnym szwem” a na ilustracjach trzony kręgów grzbietowych i ich wyrostki są w większości dysartykułowane (wyjątkiem jest kręg „f”). Wyraźnie widoczne są jednak szwy między kręgami krzyżowymi, których obecność u bazalnych (niecelurozaurowych) tetanurów sugeruje niedojrzałość (Brusatte i in., 2008, s.33; Rauhut, 2005).

Ibrahim i in. (2014) szacują, że FSAC-KK 11888 - ok. 17-letni, nie w pełni wyrośnięty osobnik - był o 32% mniejszy niż największe okazy (tj. 15-metrowe), miałby więc ok. 10,2 m. Nie zgadza się to jednak z danymi podanymi w suplemencie. W tab. S1 podano, że długość od końca pyska do tylnego końca miednicy wynosi u niego 5,88 m, więc na ogon pozostałoby 4,32 m (42% całkowitej długości). Jednak na rekonstrukcji szkieletu (Ibrahim i in., 2014, rys. F3) ogon stanowi ok. 48% długości ciała. Ibrahim (prywatna korespondencja do MZ, 2015) wyjaśnia, że rekonstrukcja nie jest „płaska”, więc mogą występować kilkuprocentowe różnice. Mortimer (online 2016) długość tego okazu oszacował na ok. 11,3 m, Cau (online 2014C) na 11 m a wg rekonstrukcji obok ma ok. 9,5 m.
BSPG 2006 I 57 – szósty kręg szyjny (piąty wg Smytha i in., 2020) – ma długość 19,8 cm (Evers i in., 2015), jest więc ok. 7% dłuższy niż piąty kręg szyjny holotypu (18,5 cm - Stromer, 1915). Jest od niego także szerszy (9,1 do ok. 7 cm), lecz niższy (8,5 do ok. 10 cm [w przedniej strony trzonu] oraz 10 do ok. 12,5 cm [w tylnej części trzonu; wysokość u holotypu oszacowana według ryc. 2 z tabl. 2 z pracy Stromera]), co potwierdza, że kręg okazu typowego jest bocznie zgnieciony (Stromer, 1915). Trudno zatem ustalić, który z okazów reprezentuje większego osobnika. Przyjęcie, że BSPG 2006 I 57 reprezentuje okaz większy o 7% od typowego wskazuje na jego długość w granicach 11,8-14,1 m.
Ibrahim i in. (2020b) wykorzystując najnowsze znaleziska i dodatkowy materiał okazu FSAC-KK 11888, oszacowali jego masę na 3,2-4,2 t, wykorzystując różne warianty gęstości tkanek. Największe osobniki ważyłyby zatem 5,4-7,1 t.
Środowisko

Spinozaury, zamieszkujące Afrykę Północną na początku późnej kredy, mogły żyć w pobliżu lasów namorzynowych, polując w zbiornikach wodnych na gady lub ryby. Obok tego wielkiego drapieżcy żyło szczególnie wiele teropodów - zob. Carcharodontosaurus#Środowisko i Spinosauridae#Paleobiologia.
Spinozaur w kulturze masowej


Spinosaurus jest dobrze znany przez wielbicieli dinozaurów z powodu wielkości, nietypowego żagla i wydłużonej krokodylej czaszki.
Spinozaur od dawna przedstawiany był w książkach popularnonaukowych o dinozaurach, lecz dopiero niedawno pojawiły się dokładniejsze informacje umożliwiające pełniejszy opis zwierzęcia. Od rekonstrukcji Lapparenta i Lavocata z 1955 roku, spinozaura wyobrażano sobie jako dużego teropoda z wysoką, silną czaszką (jak u innych wielkich teropodów) i charakterystycznym grzbietowym żaglem, czasami w pozycji czworonożnej. Później, w związku z nowymi odkryciami, zrewidowano kształt łba. Zarówno starsze, jak i wiele nowych wizerunków spinozaura często nie oddająle jego rzeczywistej budowy, zniekształcając np. kształt żuchwy czy wyrostków kręgów, które zilustrowano ponad sto lat temu.
Spinozaur wystąpił w filmie Park Jurajski III z 2001 roku, w którym to przedstawiono go jako większego i silniejszego niż Tyrannosaurus. W jednej z scen spinozaur po długiej walce pokonuje tyranozaura, wykręcając mu kark. W rzeczywistości tego typu pojedynek nie mógł się wydarzyć, ponieważ teropody te dzieliło nie tylko odległe miejsce występowania ale i kilkadziesiąt milionów lat. Scena ta wywołała niemałe kontrowersje wśród fanów tyranozaura.
"Kolczasty jaszczur" wystąpił również w grach Jurassic Park: Operation Genesis i Zoo Tycoon: Dinosaur Digs, w których także był dominującym drapieżcą,
Oprócz tego Spinosaurus został przedstawiony w kilku programach popularnonaukowych. W produkcji Mega Beasts (polski tytuł Prehistoryczne Bestie), wyprodukowanym przez Discovery Channel w 2009 r. poświęcono mu w całości jeden z odcinków, zatytułowany Biggest Killer Dino. Ukazano tam spinozaura jako czołowego drapieżcę swoich czasów i niekwestionowanego władcę jego świata, z którym nie mógł się mierzyć Carcharodontosaurus ani żadne inne zwierzę. Bardziej stonowany wizerunek omawianego teropoda przedstawiono w serialu BBC Planet Dinosaur z 2011 r. Odcinek zatytułowany New Giants przedstawia Północną Afrykę sprzed 95 mln lat. Spinosaurus został tam ukazany jako wyspecjalizowany rybożerca, unikający konkurencji z karcharodontozaurem dzięki zajmowaniu odmiennej niszy ekologicznej. Mimo to w warunkach braku pożywienia stacza z nim zaciętą, ale zwycięską walkę, okupioną jednak poważnymi ranami. Kolejnym programem dokumentalnym poświęconym spinozaurowi była produkcja zatytułowana „Bestia”. Opowiada ona o przełomowym odkryciu Ibrahima i współpracowników. Po raz pierwszy zaprezentowano obraz wodnego, czworonożnego rybożercy.
Oprócz rekonstrukcji, starających się wiarygodnie odtworzyć wygląd, środowisko i zachowanie zwierzęcia, spinozaur jest także tematem prac żartobliwych, jak np. odnoszących się do popularnych memów internetowych czy Godzilli.
Spis gatunków
| Spinosaurus | Stromer, 1915 | |
| S. aegyptiacus | Stromer, 1915 | |
| S. maroccanus | Russell, 1996 | = Sigilmassasaurus brevicollis |
Galeria

Rekonstrukcja. Autor: Joschua Knüppe [1].

Rekonstrukcja (na drugim planie niepoprawnie przedstawiono roślinożerne Ouranosaurus, które żyły na tych terenach wcześniej). Autor: ABelov2014 [2].

Rekonstrukcja. Autor: RJ Palmer (arvalis) [3].

Rekonstrukcja płynącego spinozaura. Autor: ДиБгд. [4].

Rekonstrukcja. Autor: Durbed [5].

Rekonstrukcja sprzed 2014 r. Autor: Tom Parker [6].

Rekonstrukcja szkieletu. Fot.: Mariomassone [7]

Rekonstrukcja szkieletu. Fot.: Mike Bowler [8]

Rekonstrukcja szkieletu. Fot.: Ryan Somma [9]

Rekonstrukcja głowy. Autor: Stephen O'Connor [10]

Rekonstrukcja łba, oparta na wyglądzie i zachowaniu żółwi jaszczurowatych. Autor: Fred Wiernum [11].

Rekonstrukcja czaszki. Fot.: Mike Bowler [12]

Rekonstrukcja czaszki. Fot.: Ryan Somma [13]

Rekonstrukcja czaszki i muskulatury. Autor: Christophe Hendrickx [14]

Rekonstrukcja czaszki. Autor: AS [15]

Kręg szyjny (C?4) NHMUK PV R 16429. Źródło: Evers i in., 2015 [16].
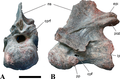
Kręg szyjny (C6-7) BSPG 2012 I 97. Źródło: Evers i in., 2015 [17].

Rekonstrukcja stopy. Fot.: Ghedoghedo [18]















![[22]](https://commons.wikimedia.org/wiki/File:Spinosaurus_aegyptiacus_holotype_skeletal.jpg){kind=link}
![[31]](https://commons.wikimedia.org/wiki/File:Spinosaurus_aegyptiacus.png){kind=link}
![[46]](https://commons.wikimedia.org/wiki/File:Spinosaurus_2020_reconstruction.jpg){kind=link}
![[4]](https://commons.wikimedia.org/wiki/File:%D0%A1%D0%BF%D0%B8%D0%BD%D0%BE%D0%B7%D0%B0%D0%B2%D1%80_-_%D0%BD%D0%BE%D0%B2%D0%B0%D1%8F_%D1%80%D0%B5%D0%BA%D0%BE%D0%BD%D1%81%D1%82%D1%80%D1%83%D0%BA%D1%86%D0%B8%D1%8F.jpg){kind=link}
![[7]](https://commons.wikimedia.org/wiki/File:Spinosaurus_white_background_2.jpg){kind=link}
![[10]](https://commons.wikimedia.org/wiki/File:Spinosaurus_skull_steveoc.jpg){kind=link}
![[14]](https://commons.wikimedia.org/wiki/File:Spinosaurus_muscles.jpg){kind=link}
![[15]](https://commons.wikimedia.org/wiki/File:Spinosaurus_skull_en.svg){kind=link}
![[18]](https://commons.wikimedia.org/wiki/File:Spinosaurus_foot_restoration.JPG){kind=link}
Bibliografia
Arden, T.M.S., Klein, C.G., Zouhri, S. & Longrich, N.R. (2018). “Aquatic adaptation in the skull of carnivorous dinosaurs (Theropoda: Spinosauridae) and the evolution of aquatic habits in spinosaurus.” Cretaceous Research. doi: https://doi.org/10.1016/j.cretres.2018.06.013.
Amiot, R., Buffetaut, E., Lécuyer, C., Wang, X., Boudad, L., Ding, Z., Fourel, F., Hutt, S., Martineau, F., Medeiros, M.A., Mo, J., Simon, L., Suteethorn, V., Sweetman, S., Tong, H., Zhang, F. & Zhou, Z. (2010) "Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods" Geology 38, 139-142.
Bailey, J.B. (1997) "Neural spine elongation in dinosaurs: Sailbacks or buffalo-backs?" Journal of Paleontology, 71(06), 1124-1146.
Beevor, T., Quigley, A., Smith, R.E., Smyth, R.S.H., Ibrahim, N., Zouhri, S. & Martill, D.M. (2020). "Taphonomic evidence supports an aquatic lifestyle for Spinosaurus." Cretaceous Research 104627 (advance online publication) doi: https://doi.org/10.1016/j.cretres.2020.104627.
Brusatte, S.L., Benson, R.B.J. & Hutt, S. (2008) "The osteology of Neovenator salerii (Dinosauria: Theropoda) from the Wealden (Barremian) of the Isle ofWight" Monograph of the Palaeontographical Society, 162(631), 1-166.
Cau, A. (online 2008) http://theropoda.blogspot.com/2008/12/misteriosi-giganti-del-sahara-quarta.html
Cau, A. (online 2009) http://theropoda.blogspot.com/2009/09/il-cranio-e-le-vertebre-dorsali-di.html
Cau, A. (online 2013A) http://theropoda.blogspot.com/2013/08/miti-e-leggende-post-moderne-sui.html
Cau, A. (online 2013B) http://theropoda.blogspot.com/2013/11/vertebre-dorsali-di-theropodi-confronto.html
Cau, A. (online 2013C) http://theropoda.blogspot.com/2013/11/loverbite-reale-ed-il-subadulto.html
Cau, A. (online 2013D) http://theropoda.blogspot.com/2013/12/ingegneria-di-un-gigante.html
Cau, A. (online 2014A) http://theropoda.blogspot.co.uk/2014/09/spinosaurus-revolution-episodio-ii-ode.html
Cau, A. (online 2014B) http://theropoda.blogspot.com/2014/09/spinosaurus-revolution-episodio-iv-una.html
Cau, A. (online 2014C) http://theropoda.blogspot.com/2014/09/spinosaurus-revolution-final-episode.html
Cau, A. (online 2015) http://theropoda.blogspot.com/2015/01/the-rise-of-tripodal-spinosaur.html
Cau, A. (online 2016A) http://theropoda.blogspot.com/2016/01/spinosaurus-war-la-parola-ai-quadrati.html
Cau, A. (online 2016B) http://theropoda.blogspot.com/2016/08/spinosaurus-gambe-corte-o-torace-lungo.html
Cau, A. (online 2016C) http://theropoda.blogspot.com/2016/08/il-canyon-la-carrucola-e-lo-spinosauro.html
Chiarenza, A.A. & Cau, A. (2016) "A large abelisaurid (Dinosauria, Theropoda) from Morocco and comments on the Cenomanian theropods from North Africa" PeerJ, 4, e1754. doi:10.7717/peerj.1754
Creisler, B. (online 2003) "Dinosauria Translation and Pronunciation Guide S" http://www.dinosauria.com/dml/names/dinos.htm [kopia: https://archive.is/e6Sto ]
Dal Sasso, C., Maganuco, S., Buffetaut, E. & Mendez, M.A. (2005) "New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities" Journal of Vertebrate Paleontology, 25(4), 888-896.
Evers, S.W., Rauhut, O.W.M., Milner, A.C., McFeeters, B. & Allain, R. (2015) "A reappraisal of the morphology and systematic position of the theropod dinosaur Sigilmassasaurus from the “middle” Cretaceous of Morocco" PeerJ, 3, e1323. doi:10.7717/peerj.1323
Fabbri, M., Navalón, G., Benson, R.B.J., Pol, D., O’Connor, J., Bhullar, B.-A.S., Erickson, G.M., Norell, M.A., Orkney, A., Lamanna, M.C., Zouhri, S., Becker, J., Emke, A., Dal Sasso, C., Bindellini, G., Maganuco, S., Auditore, M. & Ibrahim, N. (2022a) "Subaqueous foraging among carnivorous dinosaurs." Nature 603, 852–857 (2022). [[51]].
Fabbri, M., Navalón, G., Benson, R.B.J., Pol, D., O’Connor, J., Bhullar, B.-A.S., Erickson, G.M., Norell, M.A., Orkney, A., Lamanna, M.C., Zouhri, S., Becker, J., Emke, A., Dal Sasso, C., Bindellini, G., Maganuco, S., Auditore, M. & Ibrahim, N. (2022b) "Sinking a giant: quantitative macroevolutionary comparative methods debunk qualitative assumptions." bioRxiv, [[52]].
Fish, F.E., Rybczynski, N., Lauder, G.V., & Duff, C.M. (2021) "The role of the tail or lack thereof in the evolution of tetrapod aquatic propulsion." Integrative and Comparative Biology, 61:398-413. [[53]]
Franoys (online 2016) http://franoys.deviantart.com/journal/A-mathematical-analysis-on-Spinosaurus-mass-635495737
Gimsa, J., Sleigh, R. & Gimsa, U., (2015) "The riddle of Spinosaurus aegyptiacus' dorsal sail" Geological Magazine, 153(3), 544-547. doi:10.1017/S0016756815000801
Grady, J.M., Enquist, B.J., Dettweiler-Robinson, E., Wright, N.A. & Smith, F.A. (2014) "Evidence for mesothermy in dinosaurs" Science 344(6189), 1268-1272. doi:10.1126/science.1253143
Hartman, S. (online 2014A) http://www.skeletaldrawing.com/home/theres-something-fishy-about-spinosaurus9112014
Hartman, S. (online 2014B) http://www.skeletaldrawing.com/home/aquatic-spinosaurus-the-authors-responsd9182014
Headden, J. (online 2010) https://qilong.wordpress.com/2010/05/16/weekly-picture-3-spinosaurus-spines/
Headden, J. (online 2014) https://qilong.wordpress.com/2014/09/12/the-outlaw-spino-saurus/
Henderson, D.M. (2018). “A buoyancy, balance and stability challenge to the hypothesis of a semi-aquatic Spinosaurus Stromer, 1915 (Dinosauria: Theropoda).” PeerJ 6:e5409Â. [[54]].
Hendrickx, C., Mateus, O. & Buffetaut, E. (2016) "Morphofunctional analysis of the quadrate of Spinosauridae (Dinosauria: Theropoda) and the presence of Spinosaurus and a second Spinosaurine taxon in the Cenomanian of North Africa" PLoS ONE, 11(1), e0144695. doi:10.1371/journal.pone.0144695
Hone, D.W.E. & Holtz, T.R.Jr. (2021) “Evaluating the ecology of Spinosaurus: Shoreline generalist or aquatic pursuit specialist?” Palaeontologia Electronica 24(1):a03 doi: https://doi.org/10.26879/1110
Holtz, T.R., Molnar, R.E. & Currie, P.J. (2004) "Basal Tetanurae" [w:] Weishampel, D.B., Dodson, P. & Osmólska, H. "The Dinosauria" Berkeley and Los Angeles: University of California Press, 71-110.
Ibrahim, N., Sereno, P.C., Dal Sasso, C., Maganuco, S., Fabbri, M., Martill, D.M., Zouhri, S., Myhrvold, N. & Iurino, D.A. (2014) "Semiaquatic adaptations in a giant predatory dinosaur" Science, 345(6204), 1613-1616. doi:10.1126/science.1258750.
Ibrahim, N., Sereno, P.C., Varricchio, D.J., Martill, D.M., Dutheil, D.B., Unwin, D.W., Baidder, L., Larsson, H.C.F., Zouhri, S. & Kaoukaya, A/ (2020a). "Geology and paleontology of the Upper Cretaceous Kem Kem Group of eastern Morocco". ZooKeys. 928: 1–216. doi:10.3897/zookeys.928.47517.
Ibrahim, N., Maganuco, S., Dal Sasso, C., Fabbri, M., Auditore, M., Bindellini, G., Martill, D.M., Zouhri, S., Mattarelli, D.A., Unwin, D.M., Wiemann, J., Bonadonna, D., Amane, A., Jakubczak, J., Joger, U., Lauder, G.V & Pierce, S.E. (2020b). "Tail-propelled aquatic locomotion in a theropod dinosaur." Nature (2020). [[55]].
Lacerda, M.B.S., Grillo, O.N. & Romano, P.S.R. (2021): Rostral morphology of Spinosauridae (Theropoda, Megalosauroidea): premaxilla shape variation and a new phylogenetic inference, Historical Biology, DOI:10.1080/08912963.2021.2000974.
Lakin, R.J & Longrich, N.R. (2018) “Juvenile spinosaurs (Theropoda: Spinosauridae) from the middle Cretaceous of Morocco and implications for spinosaur ecology.” Cretaceous Research (in press). [56]
Maganuco, S. & Dal Sasso, C. (2018. "The smallest biggest theropod dinosaur: a tiny pedal ungual of a juvenile Spinosaurus from the Cretaceous of Morocco". PeerJ 6:e4785. doi: https://doi.org/10.7717/peerj.4785.
McFeeters, B., Ryan, M.J., Hinic-Frlog, S., Schröder-Adams, C. & Sues, H. (2013) "A reevaluation of Sigilmassasaurus brevicollis (Dinosauria) from the Cretaceous of Morocco" Canadian Journal of Earth Sciences, 50(6), 636-649. doi:10.1139/cjes-2012-0129
Mortimer, M. (online 2014) http://theropoddatabase.blogspot.com/2014/09/spinosaurus-surprise.html
Mortimer, M. (online 2016) http://theropoddatabase.com/Megalosauroidea.htm#Spinosaurusaegyptiacus
Myhrvold, N., Sereno, P.C., Baumgart, S.L., Formoso, K.K., Vidal, D., Fish, F.E. & Henderson, D.E. (2022) "Spinosaurids as “subaqueous foragers” undermined by selective sampling and problematic statistical inference.". bioRxiv doi: https://doi.org/10.1101/2022.04.13.487781
Myhrvold, N.P., Baumgart. S.L., Vidal, D., Fish, F.E., Henderson, D.M. & Saitta, E.T. (2024) "Diving dinosaurs? Caveats on the use of bone compactness and pFDA for inferring lifestyle." PLoS ONE 19(3): e0298957. [[57]]
Nash, D. (online 2014A) http://antediluviansalad.blogspot.com/2014/08/did-bakker-get-spinosaurus-right-after.html
Nash, D. (online 2014B) http://antediluviansalad.blogspot.com/2014/08/surf-or-turf-can-terrestrialaquatic.html
Nash, D. (online 2016) http://antediluviansalad.blogspot.com/2016/03/spinosaurus-unauthorized-iii-run-spino.html
National Geographic (online 2014) http://ngm.nationalgeographic.com/2014/10/spinosaurus/puzzle-graphic
Persons IV, W. S., & Currie, P. J. (2016) "An approach to scoring cursorial limb proportions in carnivorous dinosaurs and an attempt to account for allometry" Scientific reports, 6, 19828. doi:10.1038/srep19828
Rauhut, O.W.M. (2003) "The interrelationships and evolution of basal theropod dinosaurs" Special Papers in Paleontology, 69, 1-214.
Rauhut O.W.M. (2005) "Osteology and relationships of a new theropod dinosaur from the Middle Jurassic of Patagonia" Palaeontology, 48(1), 87-110.
Russell, D.A. (1996) "Isolated Dinosaur bones from the Middle Cretaceous of the Tafilalt, Morocco" Bulletin du Museum National d’histoire Naturelle. Section C, Sciences de la terre, Paleontologie,Geologie, Mineralogie, 18, 349-402.
Sampson, S.D. (2009) "Dinosaur Odyssey: Fossil Threads in the Web of Life" wyd. Princeton University, Berkeley-Los Angeles-London.
Sereno, P.C., Beck, A.L., Dutheil, D.B., Gado, B., Larsson, H.C.E., Lyon, G.H., Marcot, J.D., Rauhut, O.W.M., Sadleir, R.W., Sidor, C.A., Varrichio, D.D., Wilson, G.P. & Wilson, J.A. (1998) "A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids" Science 282, 1298-1302. doi:10.1126/science.282.5392.1298
Sereno, P.C., Fish, F.E. & Myhrvold, N. (2015) "Swimming function in the Cretaceous giant Spinosaurus aegyptiacus based on the kinematics of undulatory swimming in the American alligator" Journal of Vertebrate Paleontology, Program and Abstracts, 153.
Sereno, P.C., Myhrvold, N., Henderson, D.M., Fish, F.E., Vidal, D., Baumgart, S.L., Keillor, T.M., Formoso, K.K. & Conroy, L.L. (2022). "Spinosaurus is not an aquatic dinosaur.". bioRxiv (preprint). doi: https://doi.org/10.1101/2022.05.25.493395
Smart, S. & Sakamoto, M. (2024). "Using linear measurements to diagnose the ecological habitat of Spinosaurus. PeerJ 12:e17544 DOI 10.7717/peerj.17544.
Smith, J.B., Lamanna, M.C., Mayr, H., & Lacovara, K.J. (2006) "New information regarding the holotype of Spinosaurus aegyptiacus Stromer, 1915" Journal of Paleontology, 80(2), 400-406.
Smith, R.E. & Martill, D.W. (2023). "An unusual dental pathology in a tooth of Spinosaurus (Dinosauria, Theropoda) from the mid-Cretaceous of Morocco. Cretaceous Research. 146. 105499, [[58]].
Smyth, R.S.H., Ibrahim, N. & Martill, D.M. (2020)/ "Sigilmassasaurus is Spinosaurus: a reappraisal of African spinosaurines." Cretaceous Research 104520 (advance online publication) doi: https://doi.org/10.1016/j.cretres.2020.104520.
Stromer, E. (1914) "Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. I. Die Topographie und Geologie der Strecke Gharaq-Baharıje nebst Ausführungen über die geologisiche Geschichte-Ägyptens" Abhandlungen der Königlich Bayerischen Akademie der Wissenschaften, Mathematisch-physikalische Klasse, 16(11), 1-78.
Stromer, E. (1915) "Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. II. Wirbeltier-Reste der Baharije-Stufe (unterstes Cenoman). 3. Das Original des Theropoden Spinosaurus aegyptiacus nov. gen., nov. spec" Abhandlungen der Königlich Bayerischen Akademie der Wissenschaften, Mathematisch-physikalische Klasse, 28, 1-32. [wraz z tłumaczeniem na angielski: R. Zanon, http://paleoglot.org/browse.cfm ]
Stromer, E. (1934) "Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. II. Wirbeltier-Reste der Baharije-Stufe (unterstes Cenoman). 13. Dinosauria" Abhandlungen der Bayerischen Akademie der Wissenschaften, Mathematisch–naturwissenschaftliche Abteilung, Neue Folge, 22, 1-79. [tłumaczenie na angielski: Sven Sachs oraz ilustracja zamieszczona na http://theropoda.blogspot.com/2014/09/spinosaurus-revolution-episodio-ii-ode.html ]
Wedel, M.J. (2007) "Aligerando a los gigantes (Lightening the giants)" ¡Fundamental!, 12, 1-84. [angielskie tłumaczenie https://www.researchgate.net/profile/Mathew_Wedel/publication/252505176_LIGHTENING_THE_GIANTS_PNEUMATIC_BONES_IN_SAUROPOD_DINOSAURS_AND_THEIR_IMPLICATIONS_FOR_MASS_ESTIMATES/links/55105ed50cf20352196a07c4.pdf]
Witton, M. (online 2014A) http://markwitton-com.blogspot.co.uk/2014/09/the-spinosaurus-reboot-sailing-in.html
Witton, M. (online 2014B) http://markwitton-com.blogspot.com/2014/09/the-spinosaurus-hindlimb-controversy.html