|
|
| Linia 2: |
Linia 2: |
| | | | |
| | = Saurolophus beta = | | = Saurolophus beta = |
| − |
| |
| − | {{DISPLAYTITLE:''Saurolophus''}}
| |
| − | To jest wstępna wersja artykułu.
| |
| − | <small>
| |
| − | {| class="wikitable" style="background-color:CornSilk"
| |
| − | | Autor:
| |
| − | |-
| |
| − | | [[Łukasz Czepiński]]
| |
| − | |}
| |
| − | </small>
| |
| | | | |
| | ---- | | ---- |
| Linia 77: |
Linia 67: |
| | |} | | |} |
| | | | |
| − | ==Wstęp==
| |
| − |
| |
| − | <s>'''Zaurolof''' (''Saurolophus'') to [[rodzaj]] późnokredowego, szeroko rozpowszechnionego [[Hadrosauridae|hadrozauryda]], którego liczne szczątki znane są z [[Ameryka Północna|Ameryki Północnej]] i [[Azja|Azji]]. Ten roślinożerny dinozaur wyróżniał się charakterystycznym wyrostkiem na czaszce, tworzonym przez przedłużenie kości nosowej, a sięgającym aż za krawędź czaszki.</s>
| |
| − |
| |
| − | ==''Saurolophus osborni''==
| |
| − |
| |
| − | [[Plik:Saurolophus osborni in situ.jpg|300px|thumb|right| Ryc. 2. Szkielet holotypowy zaurolofa w trakcie wydobywania ze skał (rok 1911). Okaz leży na prawym boku, czaszka jest otoczona piaskowcem z [[ripplemarki|ripplemarkami]] (zmarszczkami), po lewej stronie znajduje się kręgosłup, zaś ogon rozciąga się aż do miejsca, w którym pracuje naukowiec. <br><br>
| |
| − | © Brown, 1913]]
| |
| − | [[Plik:Saurolophus osborni czaszka.jpg|300px|thumb|right| Ryc. 3. Czaszka ''S. osborni'' {{Kpt|Brown, 1912}} z ułamanym wyrostkiem naczaszkowym ("grzebieniem").
| |
| − | <br><br>
| |
| − | © Brown, 1912]]
| |
| − | [[Plik:Saurolophus osborni holotyp.jpg|300px|thumb|right| Ryc. 4. Holotyp ''S. osborni'' {{Kpt|Brown, 1912}} z ułamanym wyrostkiem naczaszkowym ("grzebieniem").
| |
| − | <br><br>
| |
| − | © Brown, 1912]]
| |
| − | ===Historia odkrycia===
| |
| − | <s>W 1911 roku w trakcie wykopalisk prowadzonych na terenie formacji [[Horseshoe Canyon]] (wówczas nazywana formacją [[Edmonton]]) w stanie Alberta w Kanadzie (patrz: mapa 1) zespół naukowców z Amerykańskiego Muzeum Historii Naturalnej, z paleontologiem [[Barnum Brown|Barnumem Brownem]] na czele, odkrył wspaniale zachowany, [[artykulacja|artykułowany]] szkielet zaurolofa, któremu nadany został numer katalogowy AMNH 5220 (ryc. 2, 4). Nieopodal tego samego miejsca odnaleziono również nieartykułowaną czaszkę (AMNH 5221), która została później ustanowiona [[paratyp]]em (ryc. 3)<ref name="Glut97"></ref>.
| |
| − |
| |
| − | W 1912 roku Brown opisał czaszkę zaurolofa <ref name="Brown1912"></ref>, zaś holotypowy szkielet został [[preparacja|wypreparowany]] i wystawiony w muzeum, by w 1913 roku doczekać się opisu w kolejnej publikacji<ref name="Brown1913"></ref>.</s>
| |
| − |
| |
| − | ===Materiał kopalny===
| |
| − | <s>Saurolophus osborni was named by Brown (1912). Its type specimen is AMNH 5220, a skeleton, and it is a 3D fossil preserving soft parts. Its type locality is Tolman Ferry, Red Deer River (AMNH), which is in a Campanian/Maastrichtian terrestrial horizon in Canada.<ref name="pdb_so">The Paleobiology Database. ''Saurolophus osborni''. http://paleodb.org/cgi-bin/bridge.pl?a=checkTaxonInfo&taxon_no=53386</ref>
| |
| − |
| |
| − | Holotyp: AMNH 5220, niemal kompletny szkielet z odcisiem skóry.
| |
| − |
| |
| − | Paratyp: AMNH 5221 - nieartykułowana czaszka.
| |
| − |
| |
| − | Plezjotyp: AMNH 5225 - kompletna k. kulszowa.</s>
| |
| − |
| |
| − | <s>===Diagnoza===
| |
| − | {{Szablon:Diagnoza}}
| |
| − |
| |
| − | S. osborni (Brown 1912b)
| |
| − | czaszka z długim, tylnim kostnym grzebieniem tworzonym przez wydlużenie kosci czołowej i nosowej;
| |
| − | k. lzowa bardzo długa, wyższy wyroste k. przedszczękowej wznosi się do tylnej granicy nozdrzy
| |
| − | promieniowa i barkowa podobnej długości
| |
| − | 8 kręgów krzyżowych
| |
| − | kulszowa kończy się wznosząca "stopa"
| |
| − | łonowa z któtkim wznoszącym się przednio ostrzem
| |
| − | biodrowa silnie łukowata, przedni wyrostek zakrzywiony w dół, cienki płat
| |
| − | czwarty krętarz k. udowej poniżej połowy trzony
| |
| − | palce II i IV krótie <ref name="Brown1912"></ref>
| |
| − |
| |
| − | Pierścień sklerotyczny z holotypowej czaszki wykazuje, że oko było znacznie mniejsze niż oczodół. Ruszeel 1940a wykazal, że płyty pierścienia sklerotycznego nie są szeregowe w jednym kierunku, ale raczej mają jeden lub więcej segmentów w których płytki nachodzą na siebie w odwrotnym kierunu niż w innym segmencie/segmentach<ref name="Russell40">Russell, L.S., 1940. "The sclerotic ring in the Hadrosauridae". Royal Ontario Museum, Toronto, Canada. 11 pp.</ref>.</s>
| |
| − |
| |
| − | ===Etymologia===
| |
| − |
| |
| − | <s>Nazwa [[rodzaj]]owa ''Saurolophus'' oznacza "grzebieniasty jaszczur" (z greckiego ''sauros'' - jaszczur i ''lophus'' - grzebień)<ref name="Brown1912">Brown, B. 1912. A crested dinosaur from the Edmonton Cretaceous. ''Bulletin of the American Museum of Natural History'' '''31''' (14): 131–136.</ref>. Epitet [[gatunek|gatunkowy]] - ''osborni'' - honoruje zasłużonego amerykańskiego paleontologa - [[Henry Fairfield Osborn|Henry'ego Fairfielda Osborna]].</s>
| |
| − |
| |
| − | ==''S. angustirostris''==
| |
| − |
| |
| − | ===Historia odkryć===
| |
| − | <s>W Ałtan Uła w części Efremov znaleziono siedem dużych szkieletów hadrozaurów, zachowanych w bliskim sąsiedztwie w twardym czerwonym piaskowcu. Miejsce zostało nazwane "Grobowcem Smoka" (''"Dragons' Tomb"''). Mongolski gatunek został nazwany Saurolophus angustirostris. Z tego miejsca znane są również odciski skóry tych zwierząt. Sfosylizowane płaty skóry były odlewami zrobionymi przez odwórocny obraz wciśniętej w podłoże przez ciężkie zwierzę. Oprócz tych szkieletów, rosyjscy naukowcy odnaleźli również trzy szkielety tarbozaura. [Dinosaurs of Falming Cliffs, p. 131-132].
| |
| − |
| |
| − |
| |
| − |
| |
| − | Duży okaz z Mongolii został wydobyty z formacji nemegt podczas Polsko-Mongolskich ekspedycji paleontologicznych. Przez Ryszarda Gradińskiego, Józefa Kazmierczaka i Jerzego Lefelda. Mierzył 12 metrów długości. S. angus to najbardziej obfity azjatycki hadrozaur, znany zarówno z Ałtan Uła jak i Tsagan hushu, i wydaje się być dominującym roślinożercą w formacji Nemegt. (K-J, polowanie na dinozaury; rpzygody w skamienialym swiecie; PP).
| |
| − |
| |
| − | W 1948 roku w osadach górnej kredy na południu pustyni Nemegt rosyjscy badacze z Sowiecko-Mongolskich Ekspedycji Paleontologicznych odnaleźli kompletny szkielet hadrozaura, należącego do rodzaju ''Saurolophus''. Okaz, któremu nadano numer PIN 551/8. przechowywany był w Muzeum Akademii Nauk ZSRR. Było to ważne odkrycie, gdyż odstarczyło nowych informacji na temat historii mezozoicznych gadów z Azji.
| |
| − |
| |
| − | Ivan A. Efremov wraz z Anatole K. Rozhdestvensky dowodził wyprawą.
| |
| − | </s>
| |
| − |
| |
| − | <s>===Materiał kopalny===
| |
| − | Saurolophus angustirostris was named by Rozhdestvensky (1952). Its type specimen is PIN 551-8, a skeleton, and it is a 3D body fossil. Its type locality is Nemegtu Mountain (SMPE), which is in a Campanian/Maastrichtian terrestrial horizon in the Nemegt Formation of Mongolia.<ref name="pdb_sa">The Paleobiology Database. ''Saurolophus angustirostris''. http://paleodb.org/cgi-bin/bridge.pl?a=checkTaxonInfo&taxon_no=64295</ref>
| |
| − |
| |
| − | Największym osobnikiem jest okaz (PIN 551/357 - partial adult skull), zaś najmniejszym - ZPAL MgD−1/159 (juvenile skull and partial skeleton).
| |
| − |
| |
| − | In the Nemegt Formation, S. angustirostris comprises approxi− mately 20% of all vertebrate fossils (Currie 2009; Faunal distribution in the Nemegt Formation (Upper Cre−
| |
| − | taceous), Mongolia.) found.
| |
| − | </s>
| |
| − |
| |
| − | ===Diagnoza===
| |
| − | {{Szablon:Diagnoza}}
| |
| − |
| |
| − | <s>-węzsza czaszka i dłuższy grzebień (Rozhdestevensky 1957)
| |
| − | Maryańska & Osmólska 1981a:
| |
| − | -krósze nozdrza
| |
| − | -krótsza i głębsza k. łzowa
| |
| − | -przednia część żuchwy wydłużona w długi wyrostek, wchodzący klinem między k. szczękową i łzową <ref name="Roz52"></ref>
| |
| − |
| |
| − |
| |
| − | Bazując na badaniach czaszi młodego osobnik s.angu m&o 1979 stwierdziły, że jak u innych ptasiomiednicsznych - dwie kości ponadoczodołwe są inkorporowane w górną granicę oczodołu czaszki. M&o 1979 zasugerowały rówmnjież, że utrata ponadoczodołowych jest obecna zarówno u prymitywncyh lambeozaurynow jak i hadrozaurynów, podczas gdy u bardziej zaawansowanych lambezoaurynów ponadoczodołwe mogły być początkowo częścią obręczy oczodołowej, ulegając fuzji z przedczołową i zaoczodołową w czasie ontogenezy.
| |
| − | Wykazały również 1981, że wysotek k. nosowej jest prawdopodobnie wydrążony, a lity jedynie w szczytowym końcu. Zasugerowały, że grzebien służył do zwiększenia powierzchni oddechowej otworu nosowego, mając tym samym funkcję regulacji termicznej. Również do pewnego stopnia utrata u hadrozaurynów stawów pomiędzy częściami czaszki, zwłaszcza pomiędzy dolną gałęzią . przedszczękwoej i szczekową, służyła jako coś w rodzaju amortyzator wstrząsów, chroniacy delikatną struktruę nozdrzy przed uszkodzeniem w trakcie żucia.(Maryanska, Osmolska, 79, 81)
| |
| − |
| |
| − | Uzupelniono opi s czaszki dinozaura z rodziny Hadrosauridae Saurolophus angustirostris
| |
| − | (gorna kreda Mongolii) i zwrocono uwage na jej zmiany wzrostowe. Przedstawiono hipoteze ,
| |
| − | ze grzebien nosowy wystepujacy na czaszkach przedstawicieli rodzaju Saurolophus zwi..kszal dodatkowo
| |
| − | powierzchnie jamy nosowej i sluzyl procesom term oregulacyjnym. Uznan o, zc przyczyna
| |
| − | wystepowania wielu luznych polaczen miedz y ko scmi czaszki dinozau row z rodziny Hadrosauridae
| |
| − | byla koniecznosc zabezpieczenia ich delikatnych i skomplikowanych stru ktur nosowych przed
| |
| − | wstrzasami powstajacyrni podczas intensywnych ruch6w zujacych szczek. Dokonano krytycznego
| |
| − | przegladu wszystkich takson6w Hadrosauridae opi san ych dotychczas z Azji
| |
| − |
| |
| − | ANATOMIA SZKIELBTU POZACZASZKOWBGO SAUROLOPHUS ANGUSTIROSTRlS
| |
| − | Streszczenie. - Praca zawiera dalszo rezuItaty badati autorek nad dlnozaurami kaczodziobymi (Hadrosaurldae). Dokonano
| |
| − | uzupelniajacego opisu szkieletu pozaczaszkowego Saurolophus angustirostrls ROZHDESTVENSKY, 1952, opierajac
| |
| − | sic< na materialach pochodzacych z forrnacji Nemegt (g6rna kreda) w Altan Ule i Pn, Nemegcie (Kotlina Nemegt, Pustynia
| |
| − | Gobi, Mongolska Republika Ludowa).
| |
| − | 120 TERESA MARYANSKA & HALSZKA OSMOLSKA
| |
| − | Zwr6cono uwage, ze szkielet S. angustirostris, a taki:e szkielety innych kaczodziobych, charakteryzuja si~ silnym
| |
| − | wentralnym nachyleniem przedniego odcinka tulowiowej czesci kregoslupa i zwiazanym ztyrn obnizeniern pasa barkowego,
| |
| − | niespotykanymi u innych dinozaur6w. Cechy te rnoga wskazywac na wystepowanie u kaczodziobych duzego wola.
| |
| − | Ciezar wypelnionego pokarmem roslinnym wola powodowal, zapewne, koniecznosc skr6cenia odcinka ciala znajdujacego
| |
| − | sie przed punktemjego podparcia wpanewce biodrowej, aby przesunac ku tylowi srodek ciezkosci. Nachylenie ku dolowi
| |
| − | przedniego odcinka kregoslupa tulowiowego powodowalo takie skr6cenie tej czesci ciala. Mialo to duze znaczenie dla
| |
| − | utrzymania r6wnowagi ciala tych wzasadzie dwunoznych dinozaur6w. Stosunkowo ciezka glowa musiala bye trzymana
| |
| − | wczasie lokomocji wg6rze, na pionowo ustawionej szyi, i zgodnie z przemieszczajacyrn sir;; srodkiem ciezkosci przesuwana
| |
| − | odpowiednio do tylu lub do przodu (fig. 5).
| |
| − | Wykazano, ze polaczenie koscl krzyzowej z biodrowo-Ionowym regionem miednicy bylo dodatkowo wzmocnione
| |
| − | przez Zebra krzyzowe u S.angustirostris, a taki:e eo najrnniej jeszcze u kilku innych kaczodziobych dinozaur6w (fig. 4).
| |
| − | </s>
| |
| − |
| |
| − | <s>===Porównanie z ''S. osborni''===
| |
| − | [bell 2011]
| |
| − | Maryańska and Osmólska (1984) listed eight cranial characters that apparently distinguished Sauro−lophus angustirostris from other hadrosaurids; however, Nor−
| |
| − | man and Sues (2000) argued that the diagnostic characters listed by Maryańska and Osmólska (1984) for S. angusti−rostris may fall into the realm of individual variation. Horner (1992) later attempted to distinguish the two species by the presence of a “frontal buttress” (posterodorsal process sensu Bell 2011) in only S. angustirostris. This feature has since been identified as a synapomorphy of the genus (Bell 2011).
| |
| − |
| |
| − |
| |
| − | Differs from S. osborni in having a skull that is at least 20% longer among the largest adults;
| |
| − | premaxilla with strongly reflected oral margin and upturned
| |
| − | premaxillary body in lateral aspect; broadly arcing anterior
| |
| − | margin of the prenarial fossa; an elongate, anteriorly directed
| |
| − | spur on the anterior process of the jugal that separates the lac−
| |
| − | rimal and maxilla, more so than in S. osborni; shallow
| |
| − | quadratojugal notch on the quadrate; and more strongly
| |
| − | bowed quadrate in lateral view. [bell 2011]
| |
| − |
| |
| − |
| |
| − | [bell 2012]
| |
| − | W 1947 roku rosyjscy paleontolodxzy odkryli Saurolophus angustirostris w Grobowcu Smoka. Niestety wskutek działalności "kłusowników", handlarzy skamieniałoścmi duża ilość okazów zaurolofa została stracna. Niemniej jednak odciski skóry zaurolofów z Dragon's Tomb znajdują się w archiwach muzeów na całym świecie.
| |
| − |
| |
| − | Podobnie, mniej bogate odciski skóry zostaly oddnalezione wraz z holotypem (AMNH 5220) i paratypem (AMNH 5221) S. osborni z górnej formacji Horseshoe Canyon. Okazy te zostały zebrane przez B. Brown and P. Kaisen w 1911. Odciski skóry z okolicy szczęk, miednicy, stopy i ogona nie zostały wspomniane w orygionalnych publikacjach. Znany jest również trzeci okaz zebrasny przez Browna, AMNH 5271, z zachowanymi odciskami wzdłuż ogona.
| |
| − |
| |
| − | Osteologicznie gatunki Saurolophus są podobne, z dużym wyrostkiem tworzonym przez kości nosowe, czołową i przedczołową. Ogólne podobieństwo wzbudzało wątpliwości niektórych naukowców co do ważności S. angustirostris.
| |
| − |
| |
| − | Zachowane odciski skóry umożliwiły więc wykazanie pewnych charakterystycznych cech w form...
| |
| − |
| |
| − |
| |
| − | Paleontolog Phill R. Bell z Wydziału Nauk Biologicznych Uniwersytetu Alberta w Kanadzie skupił się na okazach reprezentujących odciski skóry dwóch gatunków Saurolophus - S. angustirostris z Mongolii (kilka okazów tego gatunku zostało udostępnionych przez Magdalenę Borsuk-Białynicką z Instytutu Paleobiologii PAN) i S. osborni z Kanady. Gatunki te zostały dosyć dobrze zbadane pod względem osteologicznym (na podstawie budowy kości), a teraz mogą być rozróżniane, bazując jedynie na anatomii tkanek miękkich – różnice są bowiem widoczne w kształcie i układzie łusek. To pierwszy raz w historii paleontologii kręgowców, kiedy delikatne struktury okrywające mogą zostać użyte do identyfikacji gatunkowej danego okazu.
| |
| − |
| |
| − | A różnic jest niemało – najbardziej spektakularne są odmienności w układzie łusek na ogonie – u mongoloskiego gatunku obecne są pionowe pasma wyraźnych łusek i płaskie, sporej wielkości guzki biegnące wzdłuż linii środkwoej grzbietu. Jak wynika z badań, architektura łusek jest zgodna w każdym stadium ontogenetycznym Saurolophus angustirostris, dzięki czemu potwierdza się wcześniejsza hipoteza, iż układ tych struktur jest charakterystyczny dla danego taksonu. Co więcej, układ łusek może w pewnym stopniu wpływać na ubarwienie zwierzęcia – być może S. osborni był cętkowany, a S. angustitrostris posiadał paski (ryc. 1).
| |
| − |
| |
| − | Autor pracy postuluje, by przyszłe opisy taksonomiczne zawierały ujednolicony opis odcisków skóry – w tym pozycję i orientację tych struktur na ciele zwierzęcia. Bell sam przedstawia terminologię łusek bazującą na ich wymiarach, kształcie i ornamentacji.
| |
| − | </s>
| |
| − |
| |
| − | <s>===Tropy===
| |
| − |
| |
| − | w 1999 Ishigaki doniósł o odryciu ponad 600 dużych (25-155 cm długości), trójpalczastych tropów z późnej kredy w Bugeen Tsav i Gurilin Tsav w zachodniej części postyni Gobi.
| |
| − | Zostały odryte w latach 1995-1998 przez Hayshsibara Museum of Natural Science-Mongolian Paleontological Center Joint Paleoncological Expedicion Team, zostały przypisane do zaurolofa, bazując na obfitości kościu należących do tego rodzaju na tym samym terenie w tym samym czasie.
| |
| − | Jak twierdzi Ishigakio, ponad 15 000 tropów dinozaurów z 11 różnych lokacji na Gobi zopstały znalezionych podczas tej ekspedycji - również ankylozaurydów i teropodów. <ref name>Ishigaki,S., 1999. Abundant Dinosaur Footprints from Upper Cretaceous of Gobi Desert, Mongolia. ''Journal of Vertebrate Paleontology'' '''19''', Supplement to Number 3, 54A.</ref>
| |
| − | </s>
| |
| − |
| |
| − | ===Etymologia===
| |
| − |
| |
| − | Jak podaje [[Anatoly Konstantinovich Rozhdestvensky]] w oryginalnej, rosyjskiej publikacji z 1952 roku, ''S. angustirostris'' oznacza "зауролоф узкомордый", czyli "zaurolof o wąskim pysku"<ref name="Roz52">Рождественский, A.K. 1952. Новый представитель утконосых динозавров из Верхнемеловых отложений монголии. ''Доклады Академии Наук СССР'' '''86''': 405-408. <br>[Rozhdestvensky, A.K. 1952. A new representative of the duck-billed dinosaurs from the Upper Cretaceous deposits of Mongolia. ''Doklady Akademii Nauk S.S.S.R.'' '''86''': 405–408.]</ref>.
| |
| − |
| |
| − | <s>==''S. morrisi''==
| |
| − |
| |
| − | ===Historia odkrycia===
| |
| − | w latach 1939 i 1940 odnaleziono dwa niekompletne szkielety hadrozaurydów, zebrane przez Chestera Stocka z Instytutu Technologii w Kalifornii w skałach formacji Moreno (Kalifornia; USA).
| |
| − | Okaz LACM/CIT 2670 to słabo zachowane czaszka i kości kończyn, zaś LACM/CIT 2852) to nieco lepiej zachowana niemal kompeltna czaszka is zkielet. W 1973 roku Morris zaklasyfikwoał je jako cf. Saurolophus.
| |
| − | Analizy filogenetyczne, które wykonał Phil R. Bell i David C. Evans wykazały, że okazy te stanowią takson siostrzany do saurolophus lub Edmontosaurus, nie należą jednak do żadnego z tych rodzajów [Bell, Evans, 2010].
| |
| − |
| |
| − | Jednak w 2012 roku Prieto-Marquez i Wagner ustanowili dla tych okazów nowy gatunek - S. morrisi.</s>
| |
| − |
| |
| − | ===Materiał kopalny===
| |
| − |
| |
| − | <s>LACM/CIT 2852, a skeleton including fragment of left and most of right premaxilla,
| |
| − | both maxillae, right jugal, right quadratojugal, partial right quadrate, right postorbital,
| |
| − | paroccipital process of right exoccipital, predentary, right- and posterior fragment of left dentary,
| |
| − | partial surangular, angular, and splenial, various cervical, dorsal, and caudal vertebrae, partial
| |
| − | right scapula, both ulnae, metatarsal III, and various manual and pedal elements.
| |
| − | Referred material: LACM/CIT 2760, fragmentary skull and postcrania consisting of posterior
| |
| − | region of skull roof (including partial frontals, parietal, squamosals, prootics, supraoccipital, and
| |
| − | fragmentary exoccipitals), possible conjoined distal nasals, both maxillae, nearly complete right
| |
| − | quadrate, left- and posterior half of right dentary, partial surangular and angular, various isolated
| |
| − | dentary teeth, left coracoid, left scapula missing distal end, left humerus, distal end of right
| |
| − | humerus, proximal regions of both ulnae and radii, fragments of both femora, proximal end of
| |
| − | 7
| |
| − | the left tibia, left metatarsals III, and various fragmentary manual and pedal elements. These
| |
| − | remains appear to represent a single individual approximately 30 per cent smaller than the</s>
| |
| − |
| |
| − | [p-m 2012]
| |
| − |
| |
| − | ===Diagnoza===
| |
| − | {{Szablon:Diagnoza}}
| |
| − |
| |
| − | <s>In addition to the autapomorphy noted above, Saurolophus morrisi differs from S.
| |
| − | osborni and S. angustirostris in two more ambiguous characters. The external narial foramen of
| |
| − | Prosaurolophus and Saurolophus is elongate and slit-like, and forms a tightly constricted, almost
| |
| − | V-shaped rostroventral terminus. While there is clear evidence that the acute anterior end of the
| |
| − | foramen S. morrisi also possesses a V-shaped rostral margin of the narial foramen, it is not clear
| |
| − | that the aperture was in any way slit-like. Taken at face value, the premaxilla and maxilla of
| |
| − | LACM/CIT 2852 as preserved suggest a much deeper opening. This may be partly due to
| |
| − | diagenetic deformation of the specimen, but it is not clear how any deformation could
| |
| − | geometrically account for all of the apparent expansion of the boney naris. We concur with Bell
| |
| − | and Evans (2010) that this likely represents the original morphology to some extent. A broad
| |
| − | narial foramen is likely ancestral for hadrosaurids, but its absence in Prosaurolophus and lack of
| |
| − | information about Kerberosaurus renders interpretation of the polarity of this character in S.
| |
| − | morrisi equivocal, and therefore, we have omitted it from the formal diagnosis.
| |
| − | 8
| |
| − | Numerous characters in LACM/CIT 2760, discussed in more detail below, evidence its
| |
| − | saurolophine affinities. In addition, when considered all together, these attributes form a
| |
| − | combination of characters that support referral of this specimen to Saurolophus. Such character
| |
| − | combination includes frontal doming (at least in subadults), long and extensive ectocranial
| |
| − | surface of the frontal, substantial anteroventral downwarping of the parietal sagittal crest,
| |
| − | intersquamosal joint that excludes the parietal from the occiput, long exoccipital roof above the
| |
| − | foramen magnum, subrectangular and anteriorly oriented supratemporal fenestra, relatively high
| |
| − | and extensive posterior surface of the squamosal, anteroposteriorly broad anterodorsal region of
| |
| − | the maxilla, nearly straight posterior margin of the quadrate, quadratojugal notch of the quadrate
| |
| − | being extremely wide and positioned ventral to the mid-length of the quadrate, broad proximal
| |
| − | constriction of the scapula, relatively short and robust ulna, dentary with relatively low angle of
| |
| − | ventral deflection, and dentition with very reduced or lack of marginal denticles.
| |
| − |
| |
| − | ===Etymologia===
| |
| − | ''S. morrisi'' honoruje paleontologa [[William J. Morris|Williama J. Morrisa]], który wsławił się badaniami nad morfologią funkcjonalną i historią ewolucyjną hadrozaurydów z wybrzeża Pacyfiku i Środkowego Zachodu Ameryki Północnej<ref name="PM12">Prieto-Márquez, A., Wagner, J.R. 2012. ''Saurolophus morrisi'', a new species of hadrosaurid dinosaur from the Late Cretaceous of the Pacific coast of North America. ''[[Acta Palaeontologica Polonica]] (w druku)''</ref>.</s>
| |
| − |
| |
| − | [[Plik:Saurolophus kryschtofovici.jpg|150px|thumb|right| Ryc. XX. Okaz holotypowy ''S. kryschtofovici'' {{Kpt|Riabinin, 1930}} - fragment kości kulszowej.
| |
| − | <br><br>
| |
| − | © Riabinin, 1930]]
| |
| − |
| |
| − | ==''S. kryschtofovici''==
| |
| − |
| |
| − |
| |
| − | W 1930 roku rosyjski paleontolog, [[Anatoly Nikolaevich Riabinin]] opisał fragment lewej kości kulszowej (dokładnie jej bliższy koniec; ryc. XX) jako nowy gatunek zaurolofa - ''S. kryschtofovici'' (najprawdopodobniej na cześć kogoś o nazwisku Kryschtofovic). Skamieniałość została znaleziona w Beilyie Kruchi, na północy Heilungchiang w [[Chiny|Chinach]]<ref name="Ria30">Riabinin, A.N. 1930. On the age and fauna of the dinosaur beds on the Amur River. ''Mémoir, Société Mineral Russia'' '''59''': 41–51.</ref>. Obecnie gatunek ten jest uznawany za ''[[nomen dubium]]'' i zazwyczaj jest łączony z ''S. angustitornis'', m.in. ze względu na bliskość ich występowania (patrz: Mapa 2)<ref name="Glut97">Glut, D.F. 1997. ''Saurolophus''. (w:) "Dinosaurs: The Encyclopedia". McFarland & Co. pp. 788–789.</ref>.
| |
| − |
| |
| − | A partial “booted” ischium from the Amur region of far Eastern Russia was designated the type of Saurolophus kryschtovici by Riabinin (1930) based on comparison with
| |
| − | the equally dubious plesiotype (AMNH 5225) of S. osborni. The plesiotype, an isolated but complete ischium from the same area as the holotype, was provisionally re−identified by Russell and Chamney (1967) as Hypacrosaurus; and S. kryschtovici is unanimously regarded as a nomen dubium. [Bell 2011, czaszka]
| |
| | | | |
| | ==Paleobiologia== | | ==Paleobiologia== |
| Linia 331: |
Linia 93: |
| | | | |
| | ==Analiza filogenetyczna== | | ==Analiza filogenetyczna== |
| − | ===Historia AMNH 5225===
| |
| − | Ponieważ holotypowemu okazowi brakowało jedynie dystalnego (czyli najbardziej odsuniętego od ciała) końca kości kulszowej (wchodzącej w skład miednicy), Barnum Brown przypisał znalezioną w okolicy, nieartykułowaną k. kulszową (AMNH 5225) jako należącą do zaurolofa. Błędna, jak się później okazało, decyzja amerykańskiego paleontologa wprowadziła zamieszanie w klasyfikacji zaurolofa. W 1914 roku Brown zaproponował dwie nowe nazwy grup dinozaurów - "Trachodontidae" (obecnie [[Hadrosauridae]]) i [[Saurolophinae]], do którego należał ''Saurolophus'', ''[[Corythosaurus]]'' i ''[[Lambeosaurus]]'', ze względu na podobieństwo w kształcie dystalnej główki kości kulszowej (określanym jako "''footed''") i, jak ponownie błędnie uważał Brown, grzebień na czaszce tworzony przez wyrostki tych samych kości<ref name="Brown14">Brown, B. 1914. ''Corythosaurus casuarius'', a new crested dinosaur from the Belly River Cretaceous, with provisional classification of the family Trachodontidae. ''Bulletin of the American Museum of Natural History'' '''33'''(35):559-564</ref>. W 1916 roku Barnum Brown opisał [[Prosaurolophus|prozaurolofa]] (''Prosaurolophus''), który, ze względu na podobieństwo w anatomii czaszki, został uznany za przodka zaurolofa i włączony do wcześniej wspomnianej grupy Saurolophinae. W osiem lat później, w 1924 roku, kanadyjski paleontolog [[William Parks|William Arthur Parks]] opisał niemal kompletny szkielet prozaurolofa (okaz ROM 787), na którym dokładnie widać odmienny kształt główki kości kulszowej ("''unfooted''")<ref>Parks, W. A. 1924. Dyoplosaurus acutosqameus, a new genus and species of armored dinosaur; and notes on a skeleton of Prosaurolophus maximus. University of Toronto Studies, Geological Series 18: 1-35.</ref>. Dopiero w 1954 roku amerykański paleontolog [[Charles Sternberg|Charles Mortram Sternberg]] stwierdził, że nie ma podstaw, by uznawać przypisany przez Browna materiał za należący do zaurolofa; jednocześnie uznał za wielce prawdopodobne, że kość kulszowa zaurolofa i prozaurolofa miały podobną budowę (czyli "''unfooted''")<ref>Sternberg, C. M. 1954. Classification of North American duck-billed dinosaurs. Journal of Paleontology 28: 382-383</ref>. Ostatecznie kłopotliwy fragment miednicy z Horseshoe Canyon został przypisany do ''[[Hypacrosaurus]]''<ref>Russell, D.A. and Chamney, T.P. 1967. Notes on the biostratigraphy of dinosaurian and microfossil faunas in the Edmonton Formation (Cretaceous), Alberta. National Museum of Canada Natural History Papers 35: 22.</ref><ref>Prieto-Márquez, A. 2008. Phylogeny and historical biogeography of hadrosaurid dinosaurs. Ph.D. dissertation. Department of Biological Science. 934 pp.</ref>.
| |
| | | | |
| | ===Obecne poglądy=== | | ===Obecne poglądy=== |
BRUDNOPIS; Projekty którymi się zajmuję.
Saurolophus beta
| Saurolophus (zaurolof)
|
| Długość:
|
do 12 m
|
| Masa:
|
3 t
|
| Miejsce występowania:
|
Kanada
(formacja Horseshoe Canyon; Alberta)
Mongolia
(formacja Nemegt; prowincja Omnogov)
USA
(formacja Kirtland; stan Nowy Meksyk)
Chiny
(Beiliyie Kruchi; Heilungchiang)
|
| Czas występowania
|
83,5-65,5 Ma
późna kreda (kampan-mastrycht)
|
| Systematyka
|
Dinosauria
Ornithischia
Ornithopoda
Iguanodontia
Hadrosauridae
Saurolophinae
|
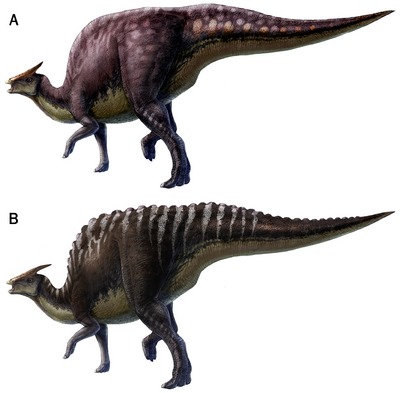
Ryc. 1. Rekonstrukcje przyżyciowe S. osborni (A) i S. angustirostris (B),
w oparciu o badania anatomii tkanek okrywających tych zwierząt z 2012 roku.
© L. Xing & Y. Liu, 2012 [1].
|
| Mapa znalezisk:
|
Wczytywanie mapy… {"minzoom":false,"maxzoom":false,"mappingservice":"leaflet","width":"auto","height":"350px","centre":false,"title":"","label":"","icon":"","visitedicon":"","lines":[],"polygons":[],"circles":[],"rectangles":[],"wmsoverlay":false,"copycoords":false,"static":false,"zoom":3,"defzoom":14,"layers":["OpenStreetMap"],"overlaylayers":[],"resizable":false,"enablefullscreen":false,"scrollwheelzoom":true,"markercluster":false,"clustermaxzoom":20,"clusterzoomonclick":true,"clustermaxradius":80,"clusterspiderfy":true,"geojson":"","locations":[],"imageoverlays":null}
Wczytywanie mapy… {"minzoom":false,"maxzoom":false,"mappingservice":"leaflet","width":"auto","height":"350px","centre":false,"title":"","label":"","icon":"","visitedicon":"","lines":[],"polygons":[],"circles":[],"rectangles":[],"wmsoverlay":false,"copycoords":false,"static":false,"zoom":3,"defzoom":14,"layers":["OpenStreetMap"],"overlaylayers":[],"resizable":false,"enablefullscreen":false,"scrollwheelzoom":true,"markercluster":false,"clustermaxzoom":20,"clusterzoomonclick":true,"clustermaxradius":80,"clusterspiderfy":true,"geojson":"","locations":[],"imageoverlays":null}
Wczytywanie mapy… {"minzoom":false,"maxzoom":false,"mappingservice":"leaflet","width":"auto","height":"350px","centre":false,"title":"","label":"","icon":"","visitedicon":"","lines":[],"polygons":[],"circles":[],"rectangles":[],"wmsoverlay":false,"copycoords":false,"static":false,"zoom":1,"defzoom":14,"layers":["OpenStreetMap"],"overlaylayers":[],"resizable":false,"enablefullscreen":false,"scrollwheelzoom":true,"markercluster":false,"clustermaxzoom":20,"clusterzoomonclick":true,"clustermaxradius":80,"clusterspiderfy":true,"geojson":"","locations":[],"imageoverlays":null} |
Paleobiologia
Saurolophus to duzy hadrozaur = holotypowysz szkielet mierzył ok. 8,4 m; ponad 2 tony [1]
Najbardziej wyróżniającą się cechą zaurolofa był kostny grzebień, który wznosi się ponad kąt czaszki niczym kolec. Brown 1912b,1913a porównywał grzebień do tego, ubecnego u kameleona, sugerując, że służył on do jako miejsce przyczepu silnych mięśni.[2][3].
Dodson 1975 - sugeruje, że grzebienie mogły mieć znaczenie w identyfikacji płciowej[4].
Paleobiogeografia
[bell 2011]
Znane są znaleziska hadrozaurynów z regionu Amur w Rosji, które zostały przypisane do blisko spokrewiononych z zaurolofem Kerberosaurus manakini. W analizie filogenetycznej kerberozaur jest prymitywniejszy niż Prosaurolophus i Saurolophus, chociaż geologicznie to Prosaurolophus jest najstarszy (kampan stanów Alberta i Montana). Saurolophus znany jest z mastrychtu formacji Horseshoe, zaś formacja Nemegt nie została dotąd dobrze orpacowana pod względem biostratygraficznym.
most lądwy Beringa pomiędzy Ameryką Północną i Azją, który byl otwarrty w trakcie aptu-albu, zapewnił rozprzestrzenianie się kręgowców lądowych w obu lokacjach w trakcie późnej kredy. Głównym kierunkiem rozprzestrzeniania się był z zachodu na wschód -
Ewolucja klady zawierającego kerberozaura prozaurolofa i zaurolofa zdradzas przynajmniej dwa zdarzenia rozptzestrzeniające pomiędzy azją a ameryką północną. Przodkowie Kerberosaurus musieli przekroczyć Azję na początku lub przed końcem kampanu. Drugie rozprzestrzenienie musiało mieć miejsce na początku mastrychtu - ze względu na kierunek pokrewieństwa pomiędzy prozaurolofem a Saurolophus osborni; prowadząc do ewolucji S. angustirostris.
Możliwe, choć mniej parsymoniczne, jest, że ostatni wspólny przodek prozaurolofa i zaurolofa przemieścił się do Azji na początku lub pod koneic kampanu, a trzecia syspersja z zachodu na wschód miała miejsce jeszcze przed końcem kampanu.
Nie jest jednak jasne który z tych dwóch gatunków jest bardziej prymitywny i w którym kierunku zachodziła dyspersja.
[prieto 2012]
Sytuację dodatkowo komplikuje wydzielenie nowego gatunku S. morrisi, którego autorzy nie pokusili się o rekonstrukcję przebiegu rozprzestrzeniania się tych zwierząt w trakcie późnej kredy. Z analizy filogenetycznej wynika, że S. morrisi jest prymitywniejszy od S. osborni i S. angustirostris.
Analiza filogenetyczna
Obecne poglądy
[bell 2011] Kerberosaurus taksonem siostrzanym do tworzonego przez Saurolophus i Prosaurolophus. Prosaurolophus siostrzany do obu gatunków Saurolophus. [prieto 2012] Saurolophus morrisi siostrzany do pozostalych gatunków zaurolofa.
prieto-marquez 2012:
Spis gatunków
Bibliografia
- ↑ Błąd rozszerzenia cite: Błąd w składni elementu
<ref>. Brak tekstu w przypisie o nazwie Glut97
- ↑ Błąd rozszerzenia cite: Błąd w składni elementu
<ref>. Brak tekstu w przypisie o nazwie Brown1912
- ↑ Brown, B. 1913. The skeleton of Saurolophus, a crested duck-billed dinosaur from the Edmonton Cretaceous. Bulletin of the American Museum of Natural History 32(19):387-393.
- ↑ Dodson, P., 1975. Taxonomic implications of relative frowth in lambeosaurine hadrosaurs. Systematic Zoology 24: 37-54.
Eskpedycje
Polsko-Mongolskie Ekspedycje Paleontologiczne
| Polsko-Mongolskie Ekspedycje Paleontologiczne
|
| Kierownictwo:
|
Zofia Kielan-Jaworowska
|
| Czas trwania:
|
1963-1965, 1967-1971
|
| Miejsce:
|
Bayn Dzak, kotlina Nemegt, Tugrig, Chermin Caw
|
| Ważne odkrycia:
|
|
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
........................
Sowiecko-Mongolskie Ekspedycje Paleontologiczne
| Sowiecko-Mongolskie Ekspedycje Paleontologiczne
|
| Kierownictwo:
|
Iwan Jefriemow
Anatol Rozdhestvensky
|
| Czas trwania:
|
1946, 1948, 1949
|
| Miejsce:
|
Bayn Dzak, kotlina Nemegt
|
| Ważne odkrycia:
|
|
W 1892 roku rosyjski odkrywca i geolog, Władimir Afanasjewicz Obruczew (Владимир Афанасьевич Обручев) opisał pierwsze skamieniałości z pustyni Gobi, zebranych w trakcie ekspedycji geograficznych pod przewodnictwem Grigorija Potanina (Григо́рий Никола́евич Пота́нин).
W latach 20-tych XX wieku Mongolia była pod dużym wpływem sowieckim, przez co Amerykkańskie Muzeum Historii Naturalnej nie mogło kontynuować swoich ekspedycji paleontologicznych po roku 1925 (zob. również Amerykańskie Ekspedycje Paleontologiczne).
w 1941 Mongolska republika ludowa zaproponowała Akademii Nauk ZSRR łączone Sowieck-Mongolskich ekspedycji paleontologicznych wgłąb Mongolii Zewnętrznej. Jednak, wskutek trwającej II Wojny Światowej, plany wspólnych ekspedycji musiały zostać przesunięte w czasie. Rozmowy zostały wznowione po zakończeniu wojny i przyniosły pożądane skutki - w 1946 roku Sowiecki Instytut Paleontologii przygotował zespół, który przeprowadził rekonesans. Zespół był dowodzony przez Iwana Jefriemowa (Ива́н Анто́нович Ефре́мов) i Anatola Konstantinovicha Rozhdestvenskyego, konsultantem był Jurij Orłow (Юрий Александрович Орлов). W skład zespołu wchodziło jeszcze sześć dodatkowych osób.
W przeciwieństwie do wcześniejszych ekspedycji, Sowieci podróżowali w dużych, ciężkich ciężarówkach i samochodach terenowych. Dzięki temu nie musieli pokładać nadziei w wielbłądzich karawanach, które dostarczały niezbędnych ładunków załodze. W ciągu dwóch miesięcy zespół sowieckich naukowców skupił się głównie na dwóch lokacjach: Jedna w połduniowowschodniej części Gobi, gdzie duże skupiska dinozaurów zostały odkryte w kotlinie Nemegt.
Większe wyprawy zostały przeprowadzone w 1948 i 1949 roku, złożone z zespołu 19 członków, 8 pracowników technicznych i 6-ciu kierowców. Dzięki szybkim samochodom, Jefriemowowi udało się zbadać głębiej położone tereny niż Amerykanom.
Pierwsze poszukiwania rozpoczęto w 1948 w lokacji Sayn Shand w południowowschodniej Gobi. Odkryto tutaj kilka szkieletów nowego ankylozauryda - Talarurus plicatospineous i fragmenatryczny materiał drugiego gatunku - Talarurus disparoserratus (obecnie zaliczanego do rodzaju Maleevus, który jest nomen dubium).
Nieco później wykopaliska przeniosły się do Bayn Dzak, gdzie Amerykanie dokonali swoich przełomowych odkryć w latach dwudziestych. Również i radzieccy uczeni znaleźli liczne szkielety Protoceratops i jaja dinozaurów. Dodatkowo odkryli dobrze zachowany szkielet dużego ankylozauryda Syrmosaurus vimicaudus (obecnie uznawany za młodszy synonim Pinacosaurus grangeri).
Gdy radziecki zespół przenioósł się w pobliże granicy z Chinami, natrafili na szerokie zagłębienie otoczoną przez pustynię. Kotlina Nemegt rozszerzała się 180 km z zachodu na wschód i ponad 70 km z północy na południe. Niebawem okazało się, że kotlina stanowi prawdziwą kopalnię dinozaurów. Osady zawierały hadrozaury, karnozaury, zaurodpoy, ankylozaury i ornitomimidy. Wzdłuż niegościnnego wąwozu czerwonego piaskowca ekspedycja odkryła siedem szkieletów Tarbosaurus bataar.
W lokalizacji niedaleko Altan Ula radzieccy naukowcy odkryli gęsty pochówek dużych hadrozaurów (ponad dwanaście metrów długości), niektóre z odciskami skóry. Samo miejsce zostało ochrzcczone nazwą "grobowiec smoka " ("Dragon's Tomb), a hadrozaury w 1952 roku przypisane do gatunku Saurolophus angustirostris.
Najbardziej zagadkowym zwierzęciem odkrytym przez swoiecko-mongolskie ekspedycje był Therizinosaurus cheloniformis, którego pazury sięgające od 30 do 60 centymettrów długości początkowo zostały uznane za żebra żółwia.
W 1949 kolejna ekspedycja podążała szlakami obranymi rok wcześniej, a również badała daleki zachód do granicy z Chinami. W Altan Teli odkryto szczątki pliocenskich ssaków. Połtorametrowej grubości warstwa zawierała szczątki tysięcy zwierząt, które mogły utonąć w niej podczas niespodziewanej powodzi. Wykopaliska zakończono powrotem do kotliny Nemegt i kontynuacją wykopalisk w miejscach, w których kopano rok wcześniej.
Radzieccy uczeni zebrali ponad 120 ton okazów paleontologicznych, w większości nieznanych dotąd nauce. Dzięki temu kotlina Nemegt, skąd pochodziła większość zebranych okazów, stała się jedną z najważniejszych lokalizacji z późnokredowymi dinozaurami, tuż obok Red Deer River w Albercie (Kanada) czy Hell Creek w Montanie (USA).
Okazy zebrane w trakcie tych ekspedycji można odlądać w Muzeum w Ułan Bator i w Muzeum Paleontologicznym Rosyjskiej Akademii Nauk w Moskwie.
2012
Magnapaulia
| Magnapaulia (magnapaulia)
|
| Długość:
|
?
|
| Masa:
|
?
|
| Miejsce występowania:
|
Meksyk
(formacja XX)
|
| Czas występowania
|
125-112 Ma
późna kreda (XX)
|
| Systematyka
|
Dinosauria
Ornithischia
Ornithopoda
Iguanodontia
Hadrosauridae
Lambeosaurinae
|
| 400px
Ryc. 1.
|
| Mapa znalezisk:
|
Wczytywanie mapy… {"minzoom":false,"maxzoom":false,"mappingservice":"leaflet","width":"auto","height":"350px","centre":false,"title":"","label":"","icon":"","visitedicon":"","lines":[],"polygons":[],"circles":[],"rectangles":[],"wmsoverlay":false,"copycoords":false,"static":false,"zoom":4,"defzoom":14,"layers":["OpenStreetMap"],"overlaylayers":[],"resizable":false,"enablefullscreen":false,"scrollwheelzoom":true,"markercluster":false,"clustermaxzoom":20,"clusterzoomonclick":true,"clustermaxradius":80,"clusterspiderfy":true,"geojson":"","locations":[],"imageoverlays":null} |
http://hadrosauria.blogspot.com/
Spis gatunków
Latirhinus
| Latirhinus (latirhin)
|
| Długość:
|
?
|
| Masa:
|
?
|
| Miejsce występowania:
|
Meksyk
(formacja XX)
|
| Czas występowania
|
83,5-70,6 Ma
późna kreda (kampan)
|
| Systematyka
|
Dinosauria
Ornithischia
Ornithopoda
Iguanodontia
Hadrosauridae
Saurolophinae
|
| 400px
Ryc. 1.
|
| Mapa znalezisk:
|
Wczytywanie mapy… {"minzoom":false,"maxzoom":false,"mappingservice":"leaflet","width":"auto","height":"350px","centre":false,"title":"","label":"","icon":"","visitedicon":"","lines":[],"polygons":[],"circles":[],"rectangles":[],"wmsoverlay":false,"copycoords":false,"static":false,"zoom":3,"defzoom":14,"layers":["OpenStreetMap"],"overlaylayers":[],"resizable":false,"enablefullscreen":false,"scrollwheelzoom":true,"markercluster":false,"clustermaxzoom":20,"clusterzoomonclick":true,"clustermaxradius":80,"clusterspiderfy":true,"geojson":"","locations":[],"imageoverlays":null} |
http://hadrosauria.blogspot.com/
Spis gatunków
Lista ootaksonów dinozaurów
- Ageroolithus
- Apheloolithus
- Arriagadoolithus
- Boletuoolithus
- Cairanoolithus
- Continuoolithus
- Dendroolithus
- Dictyoolithus
- Dispersituberoolithus
- Dughioolithus
- Ellipsoolithus
- Elongatoolithus
- Faveoolithus
- Heishanoolithus
- Laevisoolithus
- Macroolithus
- Macroelongatoolithus
- Megaloolithus
- Nanshiungoolithus
- Oblongoolithus
- Ovaloolithus
- Paraspheroolithus
- Phaceloolithus
- Placoolithus
- Porituberoolithus
- Preprismatoolithus
- Prismatoolithus
- Protoceratopsidovum
- Pseudogeckoolithus
- Shixingoolithus
- Sphaerovum
- Spheroolithus
- Spheruprismatoolithus
- Spongioolithus
- Stromatoolithus
- Subtiliolithus
- Tacumarembovum
- Trachoolithus
- Tristraguloolithus
- Youngoolithus
