Triceratops
Spis treści
- 1 Wstęp
- 2 Budowa
- 3 Budowa i paleobiologia
- 4 Filogeneza
- 5 Ontogeneza
- 6 Taksonomia - historia badań
- 7 Anageneza i heterochronia
- 8 Diagnoza
- 9 Ważność
- 10 Potencjalne synonimy
- 11 Materiał kopalny ogólnie
- 12 Lokalizacje ogólnie
- 13 T. horridus
- 14 T. sp. nov.
- 15 T. prorsus
- 16 T. sp.
- 17 ?T. sp.
- 18 Materiał wykluczony
- 19 Wątpliwe gatunki
- 20 Media
- 21 Spis gatunków
- 22 Bibliografia
- 23 Skróty instytucjonalne
| Autor: | Maciej Ziegler, Marcin Szermański |
| Triceratops (triceratops) | |
|---|---|
| Długość | 8-9 m |
| Masa | 5-13,5 (?-19)[1]t |
| Dieta | roślinożerny |
| Miejsce | USA, Kanada - zob. Lokalizacje ogólnie |
| Czas |
67[2]-66 Ma |
| Systematyka | Dinosauria |
 Rekonstrukcja T. sp.. Autor: "Laelaps nipponensis"/"GetAwayTrike". | |
Wstęp
Triceratops to rodzaj wielkiego, zaawansowanego chasmozauryna z końca kredy Ameryki Północnej.
Triceratops został opisany pod koniec XIX wieku i jest pierwszym nazwanym ceratopsem, który jest uniwersalnie uważany za ważny rodzaj (Dodson, 2013). Jest dość dobrze poznanym i jednym z najsłynniejszych mezozoicznych dinozaurów. Wiele danych na jego temat to jednak kwestia odkryć z kilkunastu ostatnich lat. Szczególnie spektakularne są badania ontogenezy, zwłaszcza nietypowe zmiany w jej trakcie (zob. też opis Torosaurus) i powiązanie ich z nowo odkrytą ewolucją w ramach tego rodzaju, a także odkrycie odcisków skóry, niepodobnych do żadnych innych dinozaurów.
Budowa
Czaszka


Czaszka ogólnie
Główne elementy budowy czaszki Triceratops są powszechnie znane (dziób, baterie zębowe, rogi nadoczne i nosowy, kryza), zob. rys. obok.
Powierzchnia czaszki
Na kryzach i rogach triceratopsów przeważnie znajdują się liczne ślady po naczyniach krwionośnych i nerwach, często tworzące wyraźną, gęstą siatkę, czasem dość szerokich i głębokich kanałów (Hatcher i in., 1907; Forster, 1996a). Taką strukturę często ma większość powierzchni czaszki (Hatcher i in., 1907) a jej obecność świadczy o pokryciu keratynową warstwą rogową (Hatcher i in., 1907, s. 21; Hieronymus i in., 2009; Horner i Marshall, 2002). Widoczne jest to szczególnie u większych osobników, lecz już u dość młodych okazów większość czaszki pokrywają takie ślady (np. UCMP 136306 o długości czaszki ok. 120 cm - Horner i Goodwin, 2014; MOR 1110 - o dł. czaszki 135 cm). Mierzące tylko 3,5 cm rogi UCMP 154452 mają takie wyżłobienia (Goodwin i in., 2006).
Podniesiono (np. Lee, online 2014), że nieopisany okaz, zachowany wraz z Nanotyrannus (znalezisko nazwane "Duelling dinosaurs") ma łuski na kryzie. Nie wiadomo jednak, jaką powierzchnię ma jego kostna kryza. Jeśli gładką, jak np. u dużego okazu UCMP 113697, to obecność łusek jest stanem, którego należy się spodziewać. Zatem znalezisko to nie podważa hipotezy o pokryciu keratyną łba Triceratops. Podobnie należy ocenić domniemany odcisk skóry u CMN 56508' (McDonald, online 2018b).
Dziób i szczęki
Triceratops był – jak zresztą każdy ceratopsyd – dinozaurem roślinożernym, wyposażonym w ostry, solidnie zbudowany i bezzębny dziób, którym skubał nisko rosnącą roślinność. Prawdopodobnie preferował rośliny zielne, choć być może przewracał na ziemię większe rośliny (jak drzewa) za pomocą rogów i dzioba (Dodson i in., 2004; s. 511). Szczęki triceratopsa wyposażone były – tak jak u innych ceratopsydów i hadrozaurydów – w baterie zębowe. U triceratopsa zaobserwowano największą ilość pozycji zębowych (36-40) spośród wszystkich przedstawicieli Ceratopsidae (Dodson i in., 2004; s. 503) (Gilmore [1920] podał 38 dla USNM 4928, Currie i in. [2016] 39 dla AMNH 970; aż 48 podał Goussard [2006, s. 472] dla MNHN 1912.20). Ich ilość zależna byłą od wielkości osobnika, np. młody UCMP 136306 miał min. 18 zębów w żuchwie (Goodwin i Horner, 2014) a UCMP 154452 ma min. 20 (Goodwin i in., 2006). W przeciwieństwie do hadrozaurydów, które mogły mielić i kruszyć pokarm roślinny, ruchy szczęk triceratopsa i jego krewnych w obrębie Ceratopsidae ograniczone były do ścierania pionowego lub prawie pionowego, były też wyjątkowo mocne (Dodson i in., 2004; s. 511). Zęby górnej i dolnej szczęki były podobnej budowy. Tylko ich górna krawędź była w użyciu (Dodson, 1996). Najwidoczniej starty ząb był zastępowany przez nowy, znajdujący się poprzednio pod nim.
Zatoki

W czaszkach ceratopsów istnieją puste przestrzenie, szczególnie rozwinięte u zaawansowanych form, takich jak Triceratops. Są to obecne u innych kręgowców (w nawiasach podano oznaczenie widoczne na rysunku obok): zatoka nosowa („nc”) i przestrzeń wypełniana przez mózg („bc”). Charakterystyczne dla ceratopsów są natomiast: zatoka czołowa (na rys.: „fs”; Farke [2010] nazywa ją „supracranial sinuses”) i będące jej częścią zatoka rogowa („cs”, „cornual diverticulum of supracranial sinus [Farke, 2010]). U Triceratops zatoka czołowa rozprzestrzenia się między kośćmi czołowymi, ciemieniowmi, zaoczodołowymi, bocznopotylicznymi (exoccipitale), nadpotylicznymi i bocznoklinowymi (Forster, 1996a). Zatoka ta miała zapewne pochodzenie pneumatyczne. Nie była połączona z zatoką nosową, w przeciwieństwie do wołowatych (Bovidae) (Farke, 2010). U niektórych osobników dostrzeżono obecność zagłębień nerwowo-naczyniowych wewnątrz zatoki rogowej (Farke, 2006). Podobieństwo zatok czołowych do znanych u wołowatych sugerowało wielu paleontologom funkcję amortyzacyjną w walkach. Dokładniejsze badania (Farke, 2008; 2010) nie dały nam pewnego wyjaśnienia roli zatok czołowych. Prawdopodobna wydaje się hipoteza, że po prostu tkanka kostna nie była potrzebna w tych miejscach (Farke, 2010).
Ciemiączka czołowo-ciemieniowe

U wielu starszych okazów Triceratops (np. YPM 1822 – zob. ryc. obok) ciemiączko ciemieniowo-czołowe (ang. frontoparietal fontanelle) i kanały grzbietowo-skroniowe (dorsotemporal channels) nie są widoczne w wyniku zarośnięcia przez kości. Nie wiadomo, czy u tych okazów pod kością znajdują się zatoki i kanały (Farke, 2010). Brown i Schlaikjer (1940, s. 163-164) sugerowali, że zamknięcie dołu czołowo-ciemieniowego (ang. frontoparietal depression) do postaci zatok (ang. supracranial sinuses) wzmacniało czaszkę, która stała się bardziej odporna na siły skierowane na rogi. Dodatkowo zamknięcie otworu tworzyło lepszą podporę dla kryzy. Kompletnie otwarty otwór w czaszce był jej słabym punktem, a początkowe jego zasklepianie - widoczne u wszystkich ceratopsydów - mogłoby służyć do wzmocnienia czaszki przeciwko obciążeniom, nawet jeżeli ciemiączko ciemieniowo-czołowe wciąż pozostało otwarte. Kompletne zamknięcie tego ciemiączka i zrośnięcie większości szwów czaszkowych mogło być kolejną adaptacją mającą na celu zmniejszenie nacisku (Farke, 2010). Potwierdza to obserwacja Mallona i in. (2011), że ciemiączko jest na ogół mniejsze u osobników Anchiceratops z długimi i mocnymi rogami nadoczodołowymi (oprócz okazu ROM 802) i większe niż u osobników z małymi rogami nadoczodołowymi. Jednak u Arrhinoceratops okaz z dłuższymi rogami ma większe ciemiączka (Mallon i in., 2014). Zamknięcie ciemiączka uznaje się za diagnostyczne dla T. prorsus - zob. komentarz do cech diagnostycznych – pkt 11, choć gatunek ten charakteryzuje się też skróconymi rogami nadocznymi i kryzą. Zamknięcie ciemiączka występuje również u charakteryzujących się krótkimi lub zresorbowanymi rogami nadocznymi przedstawicieli Centrosaurinae (Centrosaurus AMNH 5351 – Sampson i in., 1997, s. 314).
U niektórych osobników (np. T. horridus, USNM 2100; Torosaurus latus, MOR 1122), tylne końce kanałów grzbietowo-skroniowych (dorsotemporal channels) nie łączą się z oknami grzbietowo-skroniowymi (dorsotemporal fenestrae) (Forster, 1996a; Farke, 2010). U niektórych okazów (zwłaszcza T. prorsus, także Arrhinoceratops brachyops, ROM 796; Torosaurus latus, ANSP 15192) widać całkowity brak tych kanałów i zamknięcie ciemiączka ciemieniowo-czołowego.
Otwór międzyczołowy (interfrontal foramen – Farke, 2010) znajduje się między przestrzenią, gdzie znajdował się mózg, a zatokami znajdującymi się nad nim (supracranial sinuses - Farke, 2010) z pewnością u jednego z osobników ?Triceratops (CMN 8528 = USNM 5740) i według wielu autorów u innych ceratopsydów (inaczej Farke, 2010, s. 1492). Sugerowano, że połączenie mózgu z otworem na górze czaszki (ciemiączko ciemieniowo-czołowe), znamionuje obecność oka ciemieniowego. Jest to zapewne błędna hipoteza, gdyż struktura nie występowała u przodków ceratopsów (Sampson i in., 1997, s. 313). W każdym razie jeśli istniał u ceratopsów taki narząd, u T. prorsus zanikł, skoro mają one zamknięte ciemiączka.
Kłykieć potyliczny
Sferyczny kłykieć potyliczny jest obecny u wszystkich Neoceratopsia (You i Dodson, 2004, s. 486). Pozwalał prawdopodobnie na szybkie ruchy głowy w odpowiednim kierunku. Najwyższe zanotowane wśród Triceratops rozmiary kłykcia to 11,9x11,2 cm u T. prorsus MOR 1625. Większe ma prawdopodobnie jedynie Torosaurus MOR 1122 – 11,7x11,8 cm (Scannella i in., 2014) oraz ROM 59426 – 11,8x11,6 (Borkovic, 2013) o niepewnej przynależności. Nie są znane wymiary u Torosaurus MOR 981 ani u szczególnie dużych Triceratops.
Kryza
Kryza triceratopsa jest – podobnie jak u innych ceratopsydów – najbardziej charakterystyczną (diagnostyczną) częścią szkieletu, zob. (diagnoza, pkt. 2). Najdłuższą kryzę miał "Big John", którego kość łuskowa ma 112 cm (D’Anastasio i in., 2022) a najszerszą AMNH 970, którego kryza ma 157,8 cm (Lull 1903). Choć kryzę określa się jako litą, w przedniej części zawsze obecne są górne otwory skroniowe (częściowo zamknięte u FMNH P12003 - Forster, 1996a, s. 251). Miała ona u różnych okazów od poniżej 0,5 o 6 cm grubości i była najgrubsza wśród ceratopsów (Scannella i Horner, 2010). Generalnie była grubsza na obrzeżach, natomiast cieńsza na przedzie (Forster, 1996a; Horner i Goodwin, 2014), co jest widoczne już u najmłodszych okazów (Goodwin i in., 2006: 5-7 mm z tyłu i 3 mm z przodu). Na brzegu kryzy jest szeroki na 15-20 cm pogrubiony pierścień, poprzecinany śladami po naczyniach krwionośnych, który przy środku kryzy rozciąga się ku przodowi (Forster, 1990, s. 49; Forster, 1996a; widoczny np. na tablicach w Hatcher i in., 1907 i na ryc. 9A w Longrich i Field, 2012). Nie ma go jednak u YPM 1821 (Forster, 1990, s. 49) i BSM 1958G00003.00001 (Lull, 1933). Przykładowo u OMNH 10170 obrzeża kryzy mają 2,5-3,5 cm grubości a przy środku ok. 1 cm (Longrich, 2011). Na brzegach kryzy wyrastają różki - episquamosales z kości łuskowej i epiparietales z kości ciemieniowej; przeważnie (Goodwin i Horner, 2014; Scannella i in., 2014) obecny jest także epiparietosquamosale na granicy tych kości. Czasem pierwsze 1-2 episquamosales wyrastają z górnej a nie bocznej powierzchni kości łuskowych (YPM 1821 i YPM 1823 - Forster, 1996a, s. 252; jest to obecne także i niektórych Torosaurus - Farke, 2006). U większych i starszych osobników cała górna powierzchnia jest pokryta siatką nerwowo-naczyniową (zob. #Powierzchnia czaszki); wyjątkami są prawdopodobnie dość młode (Longrich i Field, 2012), lecz duże okazy AMNH 5116 (Scannella i Horner, 2010) (zob. też Torosaurus: okna), YPM 1821 (Hatcher i in., 1907, s. 145 i tablice; Longrich i Field, 2012), UCMP 113697 i USNM 1201 (Longrich i Field, 2012). Ta siatka rozciąga się ona także na dolnej powierzchni kryzy, na szerokiej na ok. 20 cm, pogrubionej jej części wokół krawędzi. Kryza łączyła się z szyją prawdopodobnie w miejscu, gdzie kończy się gruba, pokryta wyżłobieniami powierzchnia (Forster, 1996a, s. 252-253) .






Czaszka T. prorsus (holotyp) w kilku rzutach. Źródła: Hatcher i in., 1907 (rysunki) oraz Longrich i Field, 2012 (zdjęcie).
Długość rogów nadocznych


Długość rogów nadocznych zależała od gatunku (zob. pkt 10 w „górne osady”), ontogenezy i zapewne też od różnic między osobnikami.
Podawane długości rogów dotyczą z oczywistych względów kostnego trzonu rogu, bez warstwy rogowej (keratynowej), która sprawiała, że za życia zwierzęcia rogi były znacznie dłuższe. Na podstawie długości rogu jarzmowego Psittacosaurus oszacowano, że długość rogu za życia to ok. 140% długości ich kostnego trzonu (wartość u osteodermów ankylozaura Borealopelta to 125%) (Bell i in., 2022).
Długość rogu zależy od sposobu pomiaru - od punktu wybranego jako podstawa i mierzenia w linii prostej lub wzdłuż kości, np. u USNM 1201 długość (w linii prostej?) od koniuszka rogu do górnego (superior) brzegu oczodołu to 74 cm, wzdłuż przedniej części rogu to 69 cm a wzdłuż tylnej jego krawędzi to 94 cm (Hatcher i in., 1907).
Nie licząc bardzo młodych okazów, najmniejsze rogi w porównaniu do wielkości czaszki mógł mieć T. prorsus MOR 2972 - 26,7 cm w porównaniu do długości podstawowej czaszki ok. 98 cm (oszacowane na podstawie kłykcia potylicznego), co daje stosunek 0,27, lecz zielony kolor czcionki użyty we wpisie dotyczącym tego wymiaru w suplemencie publikacji Scannelli i in. (2014) w części dotyczącej rogów nadocznych oznacza zapewne to samo, co w części dotyczącej rogów nosowych, tj. zniekształcenie tafonomiczne (Scannella i in., 2014). Dłuższe rogi miał T. prorsus UCMP 136092 (Goodwin i Horner, 2010, ryc. 36.11; Scannella i in., 2014) – mają one 44 cm w porównaniu do długości podstawowej czaszki 112 cm (stosunek długości to 0,39). Zob. też opis Tatankaceratops. AMNH 5030 – okaz o niepewnej identyfikacji miał je podobnej długości, lecz proporcjonalnie nieco krótsze; długością podstawowa czaszki to ok. 117 cm (oszacowane na podstawie kłykcia potylicznego, dane z Anderson, 1999 i Farke, online 2012b) a stosunek długości to 0,38.
U AMNH 5116 długość rogu od oczodołu to 83 cm (Osborn, 1933); są to rogi proste i cienkie. U UCMP 113697 długość rogu nad oczodołem to 82 cm (Horner i Goodwin, 2006) albo 83,6 cm (Scannella i in., 2014). Największe dobrze udokumentowane rogi ma okaz MOR 3027 - 90 cm, przy czym nie są one kompletne i okaz nie wyróżnia się szczególnymi rozmiarami (długość podstawowa czaszki 110 cm - Scannella i in., 2014). Oszacowana na podstawie rekonstrukcji (Scannella i in., 2014, ryc. S1A) długość kompletnego rogu to 115 cm. Rogi MOR 3027 są najdłuższe także proporcjonalnie - stosunek ich długości do długości podstawowej czaszki od 0,82 (przy niekompletnym rogu - Scannella i in., 2014) do 1,05 (przy kompletnym rogu). To znacznie więcej niż wartość u drugiego co do proporcjonalnej długości rogów AMNH 5116 - 0,76 (dane za: Forster, 1996b; Osborn, 1933; Borkovic, 2013). Szacowana długość rogów za życia u MOR 3027 to 160 cm (Bell i in., 2022) (144 cm posiłkując się osteodermami Borealopelta).
Podawane czasem długości rogów Triceratops, jak 90 cm czy ponad 1 metr (np. Horner i Goodwin, 2008, s. 142), są prawdopodobnie zaokrąglone lub przesadzone. Rogi YPM 1821 według Marsha (1889b) miały mieć długość ok. 3 stóp (=91,4 cm) (zob. omówienie YPM 1821). Figgins (1932 - w: Lull, 1933, s. 15) podał informację o czaszce (z fragmentami kryzy) z rogami mierzącymi "nie mniej niż 3 stopy" (=91,4 cm), pochodzącej z osadów z Południowej Dakoty, z których znane są Triceratops i Torosaurus ("formacja Lance" - Lull, s. 14, tą nazwą określano warstwy obecnie przypisywane do formacji Hell Creek). Rogi okazu USNM 8059, z równowiekowych osadów z Wyoming, mają ok. 91 cm, lecz nie jest pewne, od którego taksonu pochodzą (Farke, online 2012b).
Inne ceratopsy miały podobnie długie rogi. U Eotriceratops zachowany róg mierzy 74 cm (Scannella i in., 2014) lub 77 cm (Wu i in., 2007), ale koniec nie zachował się i mierzyć mógł zdaniem Wu i in. (2007) ok. 80 cm, lecz wydaje się, że jednak znacznie więcej, gdyż zachowany koniec jest szeroki; zdaniem Scannella i in. (2014) mierzyłby 85 cm. Róg Torosaurus MOR 981 mierzy (prawdopodobnie w linii prostej, jak zaznaczono na rysunku w pracy Farkego [2006]) 85,3 cm (Farke, 2006) lub 83,5 cm (Scannella i in., 2014), przy czym widoczne są wyraźne ślady jego resorpcji (Scannella i Horner, 2010, s. 1165). U holotypu Titanoceratops rogi mają 90-91 cm (linia prosta) lub 106 cm (wzdłuż kości) (Lehman, 1998; Longrich, 2011). Prawdopodobnie najdłuższy róg pochodzi od nienazwanego chasmozauryna z wczesnego mastrychtu Meksyku. Niekompletny, zachowany w czterech kawałkach i bardzo smukły róg mierzy w linii prostej ok. 95,2 cm (Porras-Múzquiz i Lehman, 2011).
Tors
Według badań okazu TCM 2001.93.1 ("Kelsey"), przedstawionych przez P. Larsona i in. na konferencji Society of Vertebrate Paleontology (2004), brzuch triceratopsa był szerszy i bardziej okrągły niż wcześniej rekonstruowany a żebra mają wklęsłe miejsca, w których mieściła się łopatka. Badania te nie zostały dotąd opublikowane.
Ogon
Jak u innych chasmozaurynów, ogon triceratopsa był dość krótki. Według niepublikowanych badań okazu TCM 2001.93.1, kręgi ogonowe były dłuższe dorsalnie niż wentralnie, przez co w widoku bocznym miały kształt podobny do klina, co mogło powodować, że ogon był krótszy niż przeważnie się przedstawia i skierowany silnie w dół (P. Larson i in., 2004).
Kończyny i lokomocja

Triceratops to – jak każdy przedstawiciel Ceratopsoidea – czworonożny roślinożerca. Kończyny na pierwszy rzut oka przypominają kończyny słoni, z wyjątkiem kości łokciowej, za sprawą jej bardzo dużego wyrostka łokciowego (Hatcher i in., 1907; s. 193). Mimo ciężkiego ciała, budowa jego śródręcza nie różniła się diametralnie od bazalnych Neoceratopsia. Przez to triceratops opierał swoje przednie kończyny głównie na trzech wewnętrznych palcach, czwarty ledwo dotykał podłoża (Garstka i Burnham, 1997; Fujiwara, 2009, s. 1144). O kopytkowatych pazurach triceratopsa wspominali już co najmniej w 1907 roku Hatcher i in. (s. 61 i 193), mimo ograniczonych ówcześnie danych na temat kości nóg.
Długo sporne było ułożenie kończyn przednich triceratopsa i innych ceratopsydów. Badania tropów z późnego mastrychtu (Lockley i Hunt, 1995) i opisanie kompletnej, artykułowanej kończyny przedniej okazu NSM PV 20379 (Fujiwara, 2009) wykazały, że kończyny były ułożone pod tułowiem, a nie rozstawione na boki (zob. też Fujiwara i Hutchinson, 2012; Thompson i Holmes, 2007 i tam cytowane). Były one ustawione stosunkowo szeroko, nieco na zewnątrz, natomiast tylne - całkowicie pionowo (Garstka i Burnham, 1997; Fujiwara, 2009), choć według niepublikowanych badań kończyny tylne były wygięte w łuk (P. Larson i in., 2004).
Skóra


Okaz HMNS PV.1506 ("Lane") zachował się z co najmniej sześcioma różnymi typami pokrycia ciała, z różnych jego części, w tym z odciskami skóry o powierzchni ponad 1 m2 (P. Larson i in., 2007). Skóra miała guzy o średnicy do 10 cm (P. Larson i in., 2007) lub 9 cm; w każdym razie są to największe stwierdzone łuski dinozaura, większe niż mierzące do 8 cm łuski ankylozaurów (Hendrickx i Bell, 2021). Według niepotwierdzonych informacji, skóra mogła być pokryta czymś w rodzaju szczeciny (Perkins i Csotonyi, 2010), lecz żaden z guzów nie wygląda na niekompletny czy złamany aby wyciągać taki wniosek.
Budowa i paleobiologia
Mózg
Hatcher i in. (1907) sugerowali małą inteligencję triceratopsa. Forster (1996a) pisze o czaszce triceratopsa (okaz SMM P62/1/1), której przestrzeń mózgowa zajmuje około 325 cm sześciennych. Sam mózg musiał być mniejszy - np. u krokodyli mózg wypełnia około 60% jamy mózgowej.
Walki wewnątrzgatunkowe

Istnieją dowody na starcia pomiędzy różnymi osobnikami w postaci ran na czaszkach triceratopsa. Symulacje pojedynków wewnątrz gatunkowych u Triceratops przeprowadzone przez Farkego (2004) wykazały różne strefy czaszki, która mogła zostać uszkodzona wskutek walk. W zależności od typu zwarcia, których Farke wyróżnił 3 (kontakt pojedynczym rogiem nadocznym, pełen kontakt i kontakt ukośny) powstawały rany na poszczególnych częściach czaszki. Uszkodzenia obejmowały odpowiednio:
- różne elementy kryzy i rogi nadoczne
- różne elementy kryzy, rogi nadoczne i k. policzkowa (jarzmowa)
- różne elementy kryzy i rogi nadoczne
Powołując się m.in. na Gilmore’a (1919) Farke (2004) pisze, że podczas walk rogi nadoczne mogły nie wytrzymywać obciążeń i ulegać złamaniom - np. okaz USNM 4708 (prawdopodobny triceratops) ma ułamany dystalny koniec rogu nadocznego. Z reguły triceratops miał być zwierzęciem stosunkowo łagodnym, a agresywny miał się stawać właśnie w okresie lęgowym (np. Hatcher i in., 1907).


Obrona przed drapieżnikami
Triceratops, będąc uzbrojony w dwa długie rogi, był dobrze przygotowany do obrony przed mięsożercami. Kryza większości ceratopsów jest raczej zbyt delikatna, by mogła służyć do obrony przed drapieżnikami. Jednak Triceratops ma ją zwartą, litą (pozbawioną okien). Jej grubość u różnych osobników w różnych miejscach waha się od 0,5 cm do ponad 6 cm (Scannella i Horner, 2010). Żaden inny ceratops nie miał tak grubej kryzy (być może poza prawdopodobnymi synonimami Triceratops, jak Nedoceratops). Rogi triceratopsa zdają się być przystosowane do obrony, gdyż u starszych i większych osobników skierowane są do przodu (Horner i Goodwin, 2006), w przeciwieństwie do np. do Einiosaurus, który nie miał rogów nadocznych, a róg nosowy był skręcony w kierunku dolnym [Sampson, 1995], przez co zapewne nie mógł tak efektywnie godzić nim w swych oponentów. Sferyczny kłykieć potyliczny pozwalał prawdopodobnie na szybkie ruchy głowy w odpowiednim kierunku.
Niektóre okazy wykazują ślady ugryzień, spowodowanych prawdopodobnie przez tyranozaura, np. SUP 9713, zaliczony do Triceratops (zob. jednak niżej) (Happ, 2008).
Termoregulacja
Dobre ukrwienie kryzy, a także sama jej wielkość mogła mieć istotne znaczenie dla regulacji termicznej. Jak piszą Barrick i in. (1998), kryza i rogi triceratopsa służyły do termoregulacji zwierzęcia. Intensywny lub obniżony dopływ krwi do kryzy prawdopodobnie obniżał lub podwyższał, przez co stabilizował temperaturę całego ciała. Według nich ukrwiona morfologia powierzchni kryzy i jej dorsalne (grzbietowe) położenie przypomina płyty Stegosaurus i grzebień synapsyda Edaphosaurus.
Akustyczna funkcja kryzy
Niepublikowany eksperyment przeprowadzony na czaszce Triceratops sp. wykazał, że wzmacniała ona dźwięki o niskiej częstotliwości (36 Hz). Z kolei dźwięki o wyższej częstotliwości (1176 Hz) były absorbowane i mniej niż 6% z nich było rejestrowanych za pomocą aparatury badawczej (Anton, 2000). Być może zatem kryza służyła jako detektor do wykrywania czających się nieopodal drapieżników i do komunikacji na duże odległości pomiędzy poszczególnymi osobnikami triceratopsa (Anton, 2000). Twierdzi on jednak – w prywatnej korespondencji z Donaldem Glutem – że potrzeba więcej badań morfologii i fizjologii kryz u ceratopsów pod kątem akustyki (Glut, 2003).
Kryza miejscem przyczepu mięśni?
W przeszłości podnoszono, że kryzy u ceratopsów, w tym u Triceratops, były miejscem zaczepienia potężnych mięśni poruszających szczękami. Jak argumentuje np. Forster (1996a, s. 251), mały rozmiar górnego okna skroniowego i ślady po naczyniach krwionośnych u Triceratops sugerują, że nie jest to prawdziwa hipoteza. Prawdopodobnie taka właśnie była pierwotna funkcja kryzy u Ceratopsia, jednak u bardziej zaawansowanych form, takich jak Triceratops, funkcja straciła na znaczeniu na rzecz np. pokazowej (Prieto-Márquez i in., 2020).
Stadny lub samotny tryb życia


Być może triceratops prowadził stadny tryb życia, tak jak wiele innych ceratopsydów, np. Einiosaurus (Sampson, 1995; Dodson i in., 2004). Gdyby triceratops rzeczywiście był stadnym dinozaurem, wówczas zapora utworzona przez dorosłe osobniki skutecznie odstraszałaby drapieżniki, w tym Tyrannosaurus rex. Dzisiaj taką skuteczną taktykę przyjmują m.in. bawoły wobec lwów.
Nie istnieją niewątpliwe dowody na stadny tryb życia triceratopsa (Keenan i Scannella, 2014). Mathews i in. (2009) piszą o złożu szczątków trzech młodych triceratopsów, które zginęły prawdopodobnie wskutek powodzi. Keenan i Scannella (2014) opisali kolejne stanowisko zawierające trzy okazy (jeden znaleziony niżej o ok. 1 metr), tym razem w różnym wieku. Według interpretacji autorów, znalezisko nie musi świadczyć o tym, że zwierzęta zginęły razem, podobnie jak w przypadku stanowiska opisanego przez Mathewsa i in. (2009). Autorzy ci wskazali, że także doniesienie w pracy Hatchera i in. (1907, s. 136) o znalezieniu dwóch okazów oddalonych o ok. 10 stóp (ok. 3 m), niekoniecznie wskazuje na jednoczesną śmierć obu osobników.
Jest kilka nieoficjalnych doniesień o odnalezieniu większej ilości okazów w jednym miejscu. Znaleziono trzy lub cztery okazy w Wyoming, w tym dwa młode i jednego starszego osobnika (Huigen, online 2013; Smith, online 2013; Switek, online 2013). W późniejszym doniesieniu, dotyczącym prawdopodobnie tego samego znaleziska, P. Larson (w Jones, online 2016) doprecyzował, że chodzi o dwa okazy dorosłe i po jednym dorastającym i młodym. Nietypowe jest odnalezienie w jednym stanowisku kości triceratopsa i kości ceratopsa, które swoją morfologią przypomina Torosaurus oraz Triceratops (Penkalski i Skulan, 2010; Keenan i Scannella, 2014; Scannella i Horner, 2010). Manning i Egerton (2014) podali, że w Dakocie Południowej (formacja Hell Creek) znaleziono złoże, na którego obrzeżach znajduje się co najmniej 12 ceratopsów. Autorzy wskazują, że odnalezienie w jednym miejscu Torosaurus i Triceratops może rzucić „krytyczne światło” na zagadnienie ich odrębności lub tożsamości, nie podają jednak wprost, czy w opisanym przez nich stanowisku taka sytuacja występuje.
Środowisko
Zob. opisy: Leptoceratops, Edmontosaurus i Anzu.
Filogeneza
Z powodu krótkiej kryzy, Triceratops aż do lat 60-tych (np. Dodson, 2013) łączony był z przedstawicielami Centrosaurinae (grupy ceratopsów z krótkimi pyskami i kryzami, jak Centrosaurus czy Styracosaurus). Obecnie uważany jest jednak za zaawansowanego chasmozauryna (Chasmosaurinae - grupy ceratopsów z długimi pyskami i kryzami, jak Chasmosaurus czy Anchiceratops).
Triceratops należy do Triceratopsini, wraz z potencjalnymi synonimami - Nedoceratops, Torosaurus, Ojoceratops, ?Torosaurus utahensis (dwa ostatnie mogą być przodkami Triceratops według niepublikowanych badań Fowlera [2010]), prawdopodobnie też Tatankaceratops (Longrich, 2014; zob. opis Tatankaceratops) oraz najbliższymi krewnymi i prawdopodobnymi przodkami - Eotriceratops i być może Titanoceratops (?=Pentaceratops, zob. też Torosaurus: Dalekie pokrewieństwo z Triceratops). Triceratopsini to grupa najbardziej zaawansowanych chasmozaurynów, wywodzących się prawdopodobnie od ceratopsów podobnych do Anchiceratops, Arrhinoceratops, Bravoceratops lub Coahuilaceratops a może od któregoś z nich (m.in. Wu i in., 2007; Sampson i in., 2010; Longrich, 2011; Wick i Lehman, 2013). Jest całkiem prawdopodobne, że Triceratops wywodzi się od Torosaurus, którego z kolei przodkiem był Eotriceratops.
Według alternatywnej hipotezy (np. Mallon i in., 2014; 2016), Chasmosaurinae wcześnie podzieliły się na Triceratopsini (grupa z krótszymi, szerokimi kryzami) i drugi klad, zawierający pozostałe chasmozauryny (z dłuższymi, ażurowymi kryzami); pozycja Anchiceratops i Arrhinoceratops jest wśród nich niepewna.
Analiza Ryana (2007) wykazała natomiast, że Triceratops jest najbardziej bazalnym chasmozaurynem, bardziej zaawansowany jest Torosaurus, który jest taksonem siostrzanym kladu ((Anchiceratops+Arrhinoceratops)(Chasmosaurus+Pentaceratops)); analiza ta ma jednak bardzo mało cech (36) i taksonów Chasmosaurinae (6) oraz nie współgra ze stratygrafią, zaś cecha łącząca Torosaurus z bardziej zaawansowanymi chasmozaurynami jest polimorficzna u Torosaurus (Scannella i Horner, 2011).
Ontogeneza

Ontogeneza Triceratops (identyfikacja większości okazów jako T. horridus jest wątpliwa, zob. wpisy poszczególnych okazach). Autor: "Laelaps nipponensis"/"GetAwayTrike".


Badania ontogenezy
Zmiany dostrzeżone w ornamentacji czaszki są wynikiem poszukiwań skamieniałości Triceratops nieskupionych – jak większość poprzednich wypraw – na zbieraniu wielkich, kompletnych czaszek (Goodwin i Horner, 2010; 2014). Opublikowane zostały przez Hornera i Goodwina (2006; 2008) oraz Goodwina i in. (2006), którzy opisali najmniejszą znaną w miarę kompletną czaszkę (UCMP 154452). Analizę numeryczną ontogenezy przedstawili Longrich i Field (2012).
Niektóre z niżej opisanych zmian były heterochroniczne, tj. występowało ich opóźnienie lub przyspieszenie w ontogenezie w trakcie ewolucji - zob. Anageneza i heterochronia niżej.
Końcowym stadium ontogenezy według Scannelli i Hornera (2010) jest forma nazywana Torosaurus (zob. opis).
Typowe zmiany
Jak to często ma miejsce wśród kręgowców, u najmłodszych okazów (UCMP 154452) część twarzowa czaszki jest krótka a oczodoły duże. Kości czaszki są prążkowane i porowate, co znamionuje szybki wzrost. Żuchwa jest początkowo płytka, lecz szybko stawała się wysoka. Kości czaszki zrastały się, co usztywniało i być może wzmacniało czaszkę. Różne kości zrastały się w różnym tempie, co stanowiło w miarę przewidywalny ciąg. Przykładowo już u najmłodszych osobników kości ciemieniowe są zrośnięte bez widocznych szwów, kości czołowe i nosowe zrastają się później a na końcu przyrasta dziób i różki na brzegu kryzy (episquamosales, począwszy od położonych z przodu a później epiparietales). Niektóre kości, jak łuskowa i ciemieniowa, nie zrastały się nawet u największych okazów (Horner i Goodwin [2006], Ostrom i Wellnhofer [1986] podają jednak, że kości te są zrośnięte u BSP 1964 I 456). Kolejność zrastania się zależała m.in. od różnic między gatunkami (zob. Anageneza i heterochronia). Przykładowo duży róg nosowy u T. prorsus przyrastał do czaszki później, niż mniejszy róg T. horridus, lecz wcześniej zarastały ciemiączka czołowo-ciemieniowe (zob. opis tej części czaszki). Różny czas zrastania wynikał też zapewne z przypadkowej zmienności wewnątrzpopulacyjnej. Zmienny czas zrastania się dzioba, kości przedszczękowych i nosowych może być przyczyną lub skutkiem różnych preferencji żywieniowych wśród osobników w różnym wieku.
Rogi nadoczne były początkowo małymi guzkami, najpierw głównie się wydłużały a później, przez poszerzanie dolnej części, stawały się masywne. Wraz ze wzrostem rogów w ich podstawy wkraczały zatoki (kostny trzon stawał się "wydrążony"). Rósł także róg nosowy. Kryza u najmniejszego osobnika mierzy około połowy długości podstawowej czaszki (tj. długości róg-kłykieć potyliczny). Kryza jest krótka, raczej płaska i prostokątna w widoku z góry. Jest ona znacznie większa już u niewiele starszych młodych. Kości łuskowe proporcjonalnie wydłużają się przez całą ontogenezę (Scannella i Horner, 2010; Scannella i in., 2014).
Ontogeneza pozaczaszkowej części szkieletu jest słabo znana, zapewne była podobna do znanej u innych kręgowców. Długa na 10 cm kość ramienna pochodząca od ceratopsyda z formacji Frenchman jest bardzo smukła w porównaniu do czterokrotnie większej kości dorosłego Triceratops (Tokaryk, 2002). Podobne zmiany dotyczyły zapewne także innych kości. Wiele kości zrastało się, m.in. łopatka z kością kruczą czy kręgi krzyżowe (Fujiwara i Takakuwa, 2011 i tam cytowane).


Nietypowe zmiany
Inne zmiany w ontogenezie Triceratops były nietypowe, obecne prawdopodobnie tylko u Triceratops i jego najbliższych krewnych.
W ciągu życia zmieniał się kierunek zwrócenia rogów nadocznych - u najmłodszych były to niewielkie guzki. Rogi długości 5-12 cm były proste a podczas dalszego wzrostu kierowały się ku tyłowi. U starszych osobników rogi zwracały się ku przodowi - najpierw dolna część, później górna, w wyniku czego u pośrednich okazów rogi skierowany były ku przodowi ale ich końce jeszcze ku tyłowi. Zakrzywione do tyłu rogi nadoczne ma również młody Arrhinoceratops (Mallon i in., 2015). Nie jest to znane u innych ceratopsów (np. Zuniceratops - Wolfe i Kirkland, 1998 - contra Horner i Goodwin, 2006), prócz być może Agujaceratops (Lehman, 2006, ryc. 13.22 - równie dobrze można ułożyć rogi w ten sposób, że widoczne jest nieznaczne wyprostowanie rogu w ontogenezie). Zjawisko jest za to widoczne wyraźnie u rogów nosowych Centrosaurus (Frederickson, 2012; Frederickson i Tumarkin-Deratzian, 2014; Brown i in., 2020), lecz nie u Styracosaurus (Brown i in., 2020), Einiosaurus (Sampson, 1995; McDonald, 2011) (Campbell i in. [2016] wskazali na Coronosaurus, cytując Ryana i Russella [2005], lecz źródło nie potwierdza tej informacji). Za zmianę nachylenia rogów odpowiada zwiększone tempo remodelowania tkanki kostnej (Horner i Goodwin, 2005). Za życia zwierzęcia u okazów z kostnymi trzonami rogów skierowanymi całkowicie ku przodowi tylne zakrzywienie było zapewne widoczne aż do czasu, kiedy starła się pokrywająca rogi keratyna (Witton, online 2017).
U najmłodszych osobników brzeg kryzy był karbowany – wypukłe miejsca odpowiadają umiejscowieniem pojawiającym się później różkom. Różki te (episquamosales i epiparietales) początkowo były ostre i trójkątne. Wraz ze wzrostem rozciągały się i spłaszczały (Horner i Goodwin, 2006; zwrócił na to uwagę także Farke [2006, cecha 10], podając, że u dorastającego okazu z TLAM są one trójkątne a zachowuje to częściowo YPM 1822). W końcu zupełnie zrastały się z kryzą, co następowało najpierw u episquamosales położonych z przodu (Forster, 1996a). Mogło to występować także u innych Ceratopsidae. Brzeg samej kryzy (bez różków) początkowo był silnie falisty, ale szybko się wyrównywał i stawał się zupełnie gładki. Podobnie zmniejszają się inne guzki lub grzbiety na kryzie. Mogła też występować przekształcenie spiczastego rogu nosowego w płaski guz oraz skracanie rogów nadocznych (zob. więcej w artykule Zmniejszanie ozdób w ontogenezie dinozaurów: Ceratopsidae).
Zmiany w ukierunkowaniu rogów i kształcie różków na brzegu kryzy tłumaczy się funkcją rozpoznawczą (komunikacyjną) tych struktur - miały one służyć do rozpoznawania osobników przez inne jako młodych, dojrzewających czy dojrzałych. Różna orientacja rogów u młodych i starszych okazów może wynikać też z przystosowania tych drugich do walk – skierowane ku przodowi rogi zapewne nadawały się do walk wewnątrzgatunowych (por. Farke, 2004; Farke i in., 2009; Krauss i in., 2010), jak i do obrony przed drapieżnikami.

Rekonstrukcja stada Triceratops. Zwróć uwagę na kształt różków na brzegach kryzy oraz rogów nadocznych. Autor: "Laelaps nipponensis"/"GetAwayTrike".
Taksonomia - historia badań

W ciągu ponad stu lat badań wyróżniono kilkanaście gatunków Triceratops (przeważnie podaje się 16; zależnie od metody liczenia to przynajmniej 12; biorąc pod uwagę T. "ingens" [nomen nudum] i wszystkie kombinacje [tj. zapisywane jako gatunki Triceratops typowe gatunki Sterrholophus, Agathaumas, Polyonax i Nedoceratops, a także "Bison" alticornis], liczba urasta do 18; kolejnych kilka gatunków - podobnie jak Ugrosaurus olsoni - może reprezentować osobniki triceratops [tj. potencjalne synonimy, wyliczone poniżej], także Eotriceratops xerinsularis może zostać uznany za gatunek Triceratops - zatem nawet 25 nazwanych gatunków może być uznawanych za należące do Triceratops). Większość z nich opisał Othniel Marsh w XIX wieku. Za cechy diagnostyczne uznawano często kształt i rozmiary rogów oraz kryzy.
Charles Gilmore (1920, s. 100-101) wskazał, że należy brać pod uwagę czynniki takie jak płeć, ontogeneza czy zmienność osobnicza, zanim uzna się, że różnice obserwowane wśród czaszek Triceratops i innych ceratopsów są istotne taksonomicznie. Gilmore sądził też, że liczba ustanowionych gatunków jest znacząco za duża. Lull (1933, s. 115) stwierdził, że "nadawanie nazwy gatunkowej każdej dość kompletnej czaszce, jak to miało miejsce, wydaje się a priori nierozsądne". Naukowcy sugerowali pewne synonimizacje, John Hatcher i in. (1907) i Lull (1933) unieważnili kilka gatunków opartych na fragmentarycznym materiale, lecz rewizji taksonomicznej dokonano jednak dopiero pod koniec XX wieku.
Lehman w pracy magisterskiej (1982 - za Forster, 1990, s. 138) i później w publikacji z 1990 r. uznał, że Triceratops to rodzaj monotypowy, zaś różnice w budowie wynikają z płci a czasem z patologii i zaawansowanego wieku osobników. Według Sloana (1985 - za Paleobiology Database) ważne są cztery gatunki: T. horridus (= T. eurycephalus, T. flabellatus, T. serratus), T. prorsus (=T. brevicornus), T. elatus (= T. albertensis, T. calicornis) oraz T. obtusus (= Nedoceratops hatcheri).
Na podstawie porównania zróżnicowania rogów triceratopsów ze współczesnymi krętorogimi, takimi jak bawół afrykański (Syncerus caffer) i bawolec (Alcelaphus buselaphus) oraz zbliżonej pozycji stratygraficznej i geograficznej (skamieniałości niemal wszystkich gatunków odkryto na niewielkim obszarze we wschodnim Wyoming - hrabstwo Niobrara), John Ostrom i Peter Wellnhofer (1986) stwierdzili, że chodzi o jeden gatunek (T. horridus). Skupili się jednak tylko na okazach typowych i wskazali, że potrzebne są dalsze, szczegółowe badania.
Catherine Forster w dysertacji doktorskiej (1990) użyła metody kladystycznej oraz morfometrycznej do rozwiązania problemu gatunków triceratopsa (użycie połączenia obu technik było pionierskie w paleontologii). Forster uznała, że większość z 14 badanych czaszek reprezentuje T. horridus, niektóre to prawdopodobny drugi gatunek - T. prorsus a okaz AMNH 5116 na tyle różni się od pozostałych, że jest z pewnością odrębnym gatunkiem Triceratops. Na konferencji Society of Vertebrate Paleontology z 1990 r. Forster zasugerowała, że ważny jest także T. eurycephalus (Olshevsky, online 2001). Wyniki badań Forster zostały oficjalnie opublikowane, w zmienionym kształcie, dopiero w 1996 r. Analiza kladystyczna pięciu cech u trzynastu czaszek oraz analiza morfometryczna trzydziestu pięciu zmiennych u czternastu osobników wykazały, że Triceratops wykazuje dwa morfotypy. Według Forster (1996b), która oparła się na stratygrafii Lulla (1915) oba typy budowy są równomiernie rozmieszczone w czasie i przestrzeni, a ponieważ jeden z nich znacznie przewyższa liczebnością drugi (13:4), Forster wykluczyła dymorfizm płciowy i uznała, że rodzaj Triceratops składa się z dwóch gatunków: T. horridus i T. prorsus (AMNH 5116 uznała za T. horridus).
Wnioski Forster (1996b) nie zostały przyjęte przez wszystkich badaczy, np. Thomas Lehman (1998) stwierdził, że ważny jest tylko T. horridus a obserwowane różnice są gradualne. Anderew Farke (1997) na podstawie danych morfometrycznych 23 osobników wyróżnił dwa gatunki: T. horridus i T. prorsus. Ten drugi miałby mieć krótsze, bardziej zakrzywione i węziej rozstawione rogi nadoczne, krótszą, węższą i niższą kryzę oraz trochę dłuższy róg nosowy; różnice między gatunkami były gradualne. Farke zauważył, że T. horridus jest częstszy w osadach formacji Lance, a T. prorsus w położonej bardziej na północ formacji Hell Creek (obserwacja aktualna również dzisiaj: zob. lokalizacje podane przy gatunkach). Farke zgodził się też z Forster (1996b), że istotny w rozróżnianiu gatunków jest kształt pyska i zasugerował, że jest on wynikiem preferencji żywieniowych: liście zachowane w osadach formacji Hell Creek są większe, co wskazywać ma na większą ilość opadów i bujniejszą roślinność w tym środowisku. Happ i Morrow (1996) przebadali 22 czaszki i stwierdzili, że kąt przecięcia się krzywych tworzących boczne powierzchnie rogów nosowych wynosi u osobników z terenów północnych 39 +/- 8 stopni (wąskie rogi) a u osobników z południa 76+/-9 stopni (szerokie rogi), zatem ważne są dwa gatunki, rozdzielone geograficznie; badania te nie zostały opublikowane. Z geograficznym zróżnicowaniem dwóch gatunków zgodził się także Gregory Paul (2010), który uznał, że T. horridus był gatunkiem bardziej południowym a w północnej części USA (?Montana) żył obok T. prorsus, który był rzadszy i jako jedyny żył także w Kanadzie. Aktualne dane potwierdzają tą obserwację (zob. "Lokalizacje" we wpisach poszczególnych gatunków). Zwłaszcza T. prorsus jest rzadki w formacji Lance a w położonych bardziej na południe w ogóle nie występuje. Powód jest niejasny - może to wynikać choćby z większej ilości czy dostępności starszego wieku skał w tych formacjach. Proponowano także, że osobniki żyjące bardziej na północ są większe (Sloan, 1976 - za Anderson, 1999), jednak było to obalane przez innych badaczy (Anderson, 1999).
Anageneza i heterochronia





Obecnie wydaje się konsensusem podział Triceratops na dwa gatunki, głównie według kryteriów przedstawionych przez Forster (1996b) (na cechy uznawane dziś za istotne w rozróżnianiu gatunków [na długości rogów jako diagnostyczne wskazywał już Lull w 1915 r. [s. 343], który do "typu" ["phylum"] T. prorsus zaliczył jego prawdopodobny synonim T. brevicornus i niepewnie T. serratus i T. horridus, zaś do drugiego "typu" – T. elatus – zaliczył jego prawdopodobny synonim T. calicornis i niepewnie T. flabellatus oraz T. "ingens"; podobne grupy wyróżniono już wcześniej [Hatcher i in., 1907; s. 172]).
Historia badań i dane niepublikowane
Pierwsze dane o ewolucji Triceratops były dostępne jako abstrakty z konferencji naukowych na kilka lat przed ich publikacją.
Według wstępnych wyników badań Scannelli i Fowlera (2009; 2014; Horner i in., 2011), dwie morfologie (gatunki) wyróżnione przez Forster (1996b) - T. horridus i T. prorsus - są rozdzielone czasowo i reprezentują transformację ewolucyjną jednej linii – anagenezę. Od 2009 roku było wiadome, że późniejszy jest gatunek z dłuższym rogiem nosowym, czyli T. prorsus (Scannella i Fowler, 2009).
Cechy charakterystyczne T. horridus i T. prorsus są częściowo wynikiem zmian heterochronicznych - opóźnień lub przyspieszeń rozwoju danych struktur w ontogenezie w trakcie ewolucji (Scannella i Fowler, 2009; Scannella, 2010; Scannella i in., 2012). Niepublikowane dane wskazują, że osobniki znajdujące się w wyższych warstwach zachowują młodocianą morfologię pyska (ang. rostrum) w późniejszym stadium ontogenezy (pedomorfoza) a młode mają cechy dorosłych z wcześniejszych warstw (Scannella, 2010; Scannella i in., 2012; 2013). Zachowanie pierwotnych cech u młodych okazów dotyczy też budowy obecnej u bardziej bazalnych, wcześniejszych chasmozaurynów Anchiceratops, Chasmosaurus czy Pentaceratops (np. grzbiet na bocznej stronie kości zębowej jest obecny u MOR 1199) (Scannella i in., 2013). Niepublikowane dane sugerują także – wbrew tym opublikowanym – że osobniki znajdujące się w wyższych warstwach miały zredukowane rogi nadoczne w późniejszym stadium ontogenezy (Scannella, 2010).
Wyniki badań Triceratops z formacji Hell Creek zostały ostatecznie opublikowane w 2014 r. przez Scannellę, Fowlera, Goodwina i Hornera. Oto wnioski (rozbudowane) z tej publikacji:
Dolne osady

Okazy z dolnych i dolnych środkowych warstw mają budowę charakterystyczną dla T. horridus. Szczątki z plezjomorficzną morfologią występują przez ok. 31 m (od Torosaurus MOR 1122 do ?T. horridus MOR 3011) a wliczając UCMP 128561 – 50 m miąższości formacji Hell Creek, zatem zasięg czasowy jest znaczny – przekracza połowę formacji. Najstarsze okazy Triceratops (MOR 1120 i MOR 2985) pochodzą z poziomu 20 m nad podstawą formacji i 3 m nad okazami Torosaurus (MOR 3081). Same T. horridus znane są z przedziału 12 m (MOR 1120 i MOR 3011) a wliczając UCMP 128561 – 30 m. Okazy wydobyte z osadów znajdujących się ok. 40 nad podstawą formacji (niediagnostyczne MOR 2982 i MOR 3058) mają budowę nieznacznie bardziej zaawansowaną (zob. niżej) (Scannella i in., 2014, ryc. 1). Okazy z dolnych i dolnych środkowych warstw mają następujące cechy [w nawiasach kwadratowych podano numer cechy w analizie binarnej Scannelli i in., 2014]:
(1) długi pysk - stosunek długości od dzioba do podpórki nosowej kości przedszczękowej (narial strut) powyżej do długości od dzioba do kłykcia potylicznego powyżej 0,28 (MOR 1120) (Scannella i in., 2014).
(2) [3] pochylony ku tyłowi wyrostek nosowy kości przedszczękowej (MOR 1120 i MOR 3011). Kąt między wyrostkiem nosowym kości przedszczękowej (ang. nasal process of the premaxilla – NPP) a jej „rozporą nosową” (ang. narial strut) (por. Scannella i in., 2014, ryc. S2) wynosi u ww. Triceratops 126-127 stopni a średnia wszystkich okazów z dolnych i dolnych środkowych warstw (z Torosaurus i niediagnostycznych) to 123 stopnie (u Torosaurus MOR 981 to tylko 104 stopnie).
(3) [14] wąski wyrostek nosowy kości przedszczękowej (MOR 1120, MOR 3011). Stosunek wysokości (mierzonej od najniższego punktu artykulacji z kością nosową) do szerokości (długości) najwęższej części wynosi u obu 1,73 a wliczając okazy Torosaurus i niediagnostyczne od 1,59 do 3,04 i średnio 1,99 (Scannella i in., 2014).
(2-3) Obie te cechy, zwłaszcza ta pierwsza, powodowały, że pysk miał budowę opisaną przez Forster (1996b, cecha 4) (Scannella i in., 2014), tj. „wydłużony, S-kształtny pysk” u T. horridus.
(4) [21] wąską tylnobrzuszną powierzchnię dolnotylnego wyrostka kości przedszczękowej (MOR 1120).
(5) [22] szeroką powierzchnię styku z kością nosową dolnotylnego wyrostka kości przedszczękowej (MOR 1120 i niediagnostyczne MOR 2982 i MOR 3005).
(6) [7] krótszy i mniej wydłużony róg nosowy (MOR 1120 i MOR 3011). Rogi ceratopsów (Triceratops, Torosaurus i niediagnostycznych) z dolnej części formacji Hell Creek mają do 15,5 cm długości i mają średnie wydłużenie 1,04 (uwzględniono UCMP 128561 – holotyp Ugrosaurus z górnych środkowych warstw i długość MOR 966 z nieznanego poziomu). Stopień wydłużenia rogu (długość/szerokość) wynosi 0,8-1,5 (wartości dla: MOR 1120 i niediagnostycznego MOR 2982; u Torosaurus są często niższe wartości, np. 0,7 u MOR 1122). Forster (1996b, cecha 5) podała, że krótkie, zwrócone do góry rogi T. horridus mają od długość od 0,1 do 0,13 długości od dzioba do kłykcia potylicznego, natomiast analogiczne wartości – wliczając Torosaurus i niediagnostyczne okazy – według danych Scannelli i in. (2014) to od 0,07 (MOR 1120) do 0,14 (niediagnostyczny MOR 2570), średnio 0,9.
(7-8) [13] guzek na tylnej powierzchni rogu nosowego (MOR 3011) i [10-11] guzek formowany przez tylną część rogu nosowego i przednią część kości nosowej (MOR 1120).
(9) krótka kość nosowa – stosunek jej długości do długości od dzioba do kłykcia potylicznego u większych okazów (druga długość ok. 1 m) poniżej 0,38 (MOR 1120).
(10) [9] krótką i łukowato wygiętą dolną krawędź górnej części kości nosowej (MOR 1120 i Torosaurus: MOR 981, MOR 1122, MOR 3005; obecne też u YPM 1820 – Scannella i in., 2014).
(11) [12] wyraźny przedniobrzuszny wyrostek kości nosowej (MOR 1120 i MOR 3011).
(12) [1] zróżnicowanej długości, przeważnie dość długie rogi nadoczne. Rogi T. horridus i T. ?sp. nov. („pośrednich” Triceratops) są przeciętnie długie, u większych okazów (ok. 1 m i więcej długości podstawowej czaszki, tj. od koniuszka dzioba do końca kłykcia potylicznego) mają około lub ponad 50 cm. Maksymalna długość rogów u T. horridus i T. ?sp. nov. to 80-90 cm. U tych pierwszych zdarzają się okazy z dość krótkimi rogami – np. holotyp. Długość rogów u większych okazów T. horridus z dolnych i dolnych środkowych warstw to 0,47-0,74 długości podstawowej czaszki, u T. ?sp. nov. 0,58-0,82. Forster (1996b) wskazała natomiast zakresy 0,69-0,75 dla T. horridus (w tym T. ?sp. nov. UCMP 113697).
(13) [4] otwarte ciemiączka ciemieniowo-czołowe (frontoparietal fontanelle - Farke, 2010) u dorosłych okazów (Forster, 1996b).
(14) [24] płaską boczną krawędź kości łuskowej.
Górne środkowe osady
Okazy pośrednie - T. ?sp. nov. – z górnych środkowych warstw formacji Hell Creek mają cechy pośrednie między starszymi T. horridus a młodszymi T. prorsus. Pochodzą z wąskiego przedziału czasowego – 2 m na wysokości 50 m od podstawy formacji (Scannella i in., 2014, ryc. 1).
- cechy wcześniejszych T. horridus: (7-8), [10, 11, 13] (11) [12] (12) [1] - długie rogi nadoczne (0,58-0,82) - okaz z najdłuższymi znanymi rogami (MOR 3027) reprezentuje to stadium a krótsze rogi ma MOR 3045, (13) [24] płaską boczną krawędź kości łuskowej.
- cechy zbliżające je do T. prorsus:
(1) krótki pysk - stosunek długości od dzioba do podpórki nosowej kości przedszczękowej (narial strut) powyżej do długości od dzioba do kłykcia potylicznego poniżej 0,28 (Scannella i in., 2014).
(3) [14] szerszy wyrostek nosowy kości przedszczękowej. Stosunek wysokości (mierzonej od najniższego punktu artykulacji z kością nosową) do szerokości (długości) najwęższej części wynosi u dwóch okazów odpowiednio 1,17 (alternatywnie 1,39 – zob. dalej) i 1,46 (Scannella i in., 2014), tj. średnio 1,28 (alternatywnie 1,42). 1,17 dotyczy MOR 3045, bez uwzględnienia występującego poza tym okazem jedynie u młodych z górnej części formacji Hell Creek (MOR 1110 i MOR 2951) dodatkowego wyrostek na górze omawianego wyrostka. Po uwzględnieniu wysokości wyrostka otrzymuje się 1,39 i 1,42 – te wartości przyjęli Scannella i in. (2014), uznając jednocześnie, że MOR 3045 to najbardziej bazalny i najwcześniejszy z okazów wykazujących szeroki wyrostek. Wizualnie proporcje nie różnią się między tym okazem a pochodzącymi z górnych osadów (ryc. S2 w Scannella i in., 2014). UCMP 113697 tutaj wykluczono, ponieważ wysokość jest niemożliwa do zmierzenia z powodu zakrycia kości (stosunek wynosi 1,16).
(4) [21] niektóre okazy (MOR 3027) mają wąską tylnobrzuszną powierzchnię dolnotylnego wyrostka kości przedszczękowej, jak u T. horridus (MOR 1120) a inne (MOR 3045) szeroką, jak u T. prorsus.
(6) [7] bardziej wydłużony róg nosowy. Długości rogów to 16-24 cm i średnie wydłużenie równe 2 (dane dotyczą dwóch okazów). Stosunek długości rogów w porównaniu do długości od dzioba do kłykcia potylicznego to 0,16 i 0,23 (średnio 0,19).
(9) dłuższą kość nosową – stosunek jej długości do długości od dzioba do kłykcia potylicznego u większych okazów (druga długość ok. 1 m) powyżej 0,38 (MOR 3027).
(10) [9] prostszą dolną krawędź górnej części kości nosowej (MOR 3027).
- cechy T. prorsus:
(2) [3] mniej pochylony (bardziej pionowy) wyrostek nosowy kości przedszczękowej. Kąt między nim a „rozporą nosową” u okazów z górnej środkowej i górnej części formacji kąt to od 137 do 142 stopni a średnio 140 stopni (Scannella i in., 2014).
(5) [22] wyraźny grzbiet na bocznej powierzchni dolnotylnego wyrostka kości przedszczękowej i zawężoną powierzchnię styku z kością nosową.
Górne osady
Okazy z górnych warstw formacji Hell Creek mają budowę charakterystyczną dla T. prorsus. Pochodzą z przedziału o miąższości co najmniej 20 m w górnej części formacji Hell Creek (Scannella i in., 2014, ryc. 1), datowej na ok. 66,4/66,25-66 Ma (Fowler, online 2016; 2017; 2020). Kilka metrów szczytu formacji nie zawiera szczątków a pomiędzy T. prorsus a T. ?sp. nov. występuje ok. 7 m przerwy w zapisie kopalnym (Scannella i in., 2014, ryc. 1). Upraszczając można uznać, że T. prorsus występował 66,3-66 Ma (Fowler, online 2016; 2017; 2020). Okazy z górnych warstw mają następujące cechy:
- cechy wcześniejszych (T. ?sp. nov.) okazów:
(1) krótki pysk - stosunek długości od dzioba do podpórki nosowej kości przedszczękowej (narial strut) powyżej do długości od dzioba do kłykcia potylicznego poniżej 0,28 (Scannella i in., 2014).
(2) [3] mniej pochylony (bardziej pionowy) wyrostek nosowy kości przedszczękowej. Kąt wynosi od 131 (młody okaz MOR 1110) przez 135 do 142 stopni a średnio 140 stopni (Scannella i in., 2014), czyli praktycznie tyle samo, ile u wcześniejszych Triceratops.
(5) [22] wyraźny grzbiet na bocznej powierzchni dolnotylnego wyrostka kości przedszczękowej i zawężona powierzchnia styku z kością nosowej.
- bardziej nasilone cechy wcześniejszych okazów:
(3) [14] szeroki wyrostek nosowy kości przedszczękowej. Stosunek wysokości do szerokości (długości) wynosi od 0,97 (MOR 2936 – okaz niekompletny i niekoniecznie należący do Triceratops), przez 1,18 (MOR 1110, okaz młody), 1,24 (MOR 2702) do 1,157 (MOR 2951), średnio 1,24 a bez młodych 1,15.
(2-3) Te cechy powodowały „krótki i zaokrąglony pysk” (Forster, 1996b, cecha 4).
(4) [21] szeroką tylnobrzuszną powierzchnię dolnotylnego wyrostka kości przedszczękowej, jak u pochodzącego z górnych środkowych osadów MOR 3045 (obecne u MOR 2924, MOR 2702 oraz u niekoniecznie należących do Triceratops MOR 2936 i MOR 2979).
(6) [7] długi i wąski róg nosowy. Poza młodymi okazami (MOR 1110 i MOR 2951), wydłużenie rogów to 2,1-2,9, średnio 2,3 (Scannella i in., 2014). Z górnych osadów formacji Hell Creek pochodzi izolowany róg MOR 2576 o długości 21,3 cm i wydłużeniu nieznacznie większemu niż u wcześniejszych - 1,56 (Scannella i in., 2014; Scannella i Fowler, 2014). Forster (1996b, cecha 5) podała, że długie, zwrócone bardziej do przodu rogi T. prorsus mają od 0,23 do 0,3 długości od dzioba do kłykcia potylicznego, natomiast analogiczne wartości – wliczając niediagnostyczne okazy – według danych Scannelli i in. (2014) to od 0,08 (młody MOR 2951) do 0,26 (np. MWC 7584), średnio 0,21 a bez uwzględnienia najmłodszych okazów od 0,21 (MOR 2574 – dość młody - Keenan i Scannella, 2014) i średnio 0,24. Z danych Scannelli i in. (2014) wynika również, że u większych okazów rogi były bardziej wydłużone.
(9) dłuższą kość nosową – stosunek jej długości do długości od dzioba do kłykcia potylicznego u większych okazów (druga długość ok. 1,1 m) powyżej 0,41.
(10) prostszą dolną krawędź górnej części kości nosowej.
- nowe cechy:
(7-8) [10, 11, 13] zredukowany (lub brak) guz na górnej powierzchni rogu nosowego i zredukowany lub nieobecny guzek formowany przez tylną część rogu nosowego i przednią część kości nosowej.
(11) [12] zredukowany przedniobrzuszny wyrostek kości nosowej.
(12) [1] zawsze krótkie rogi nadoczne - 0,27-0,64 długości podstawowej czaszki, tj. od dzioba do kłykcia potylicznego (ta ostatnia liczba posłużyła jako wyznacznik do kwalifikowania okazów jako mających długie lub krótkie rogi w analizie kladystycznej Scannelli i in., 2014). U wielu okazów były krótsze niż 0,45 długości podstawowej czaszki (u większych okazów o tej długości większej niż 50-90 cm), czego nie odnotowuje się u wcześniejszych Triceratops (obecne też u Torosaurus latus). Rogi T. prorsus są przeciętnie krótsze i u większych okazów osiągają przeważnie 40-60 cm. Maksymalna długość rogów to ok. 60 cm (Scannella i in., 2014); u EM P15.1 ok. 65 cm (Tokaryk, 1986; zmniejszono wymiar podany przez autora, gdyż dotyczył odległości do środka oczodołu). Rogi T. prorsus są według Borkovica (2013, np. ryc. 5.21) grubsze (szersze) niż u T. horridus, lecz porównanie ich szerokości z długością podstawową czaszki (dziób-kłykieć potyliczny) wskazuje, że jest to jedynie wynikiem skrócenia rogów (ten pierwszy wymiar stanowi 8% i 15% drugiego wymiaru odpowiednio u T. horridus ROM 55380 i AMNH 5116 oraz 13% i 14% odpowiednio u T. prorsus YPM 1822 i CM 1219; u YPM 1821 to 14% a YPM 1823 – 13%). Szczególnie długie rogi jak na T. prorsus ma dość młody okaz MOR 1110 (50 cm, stosunek do długości podstawowej czaszki to 0,64). Prawdopodobnie jest to jedyny okaz z tego stadium ontogenezy (Longrich i Field, 2012) i tych rozmiarów (długość podstawowa czaszki 77,6 cm - Scannella i in., 2014; długość całkowita czaszki 135 cm - Horner i Goodwin, 2006). Jedynym zbliżonym wielkością okazem jest problematyczny MCZ 1102 (holotyp T. eurycephalus) o długości czaszki ok. 138,6 cm (Schlaikjer, 1935) i jeszcze dłuższych rogach (63 cm – Dodson, 1996, s. 78), które są zwrócone do przodu, co sugeruje, że okaz jest dojrzalszy od MOR 1110. Długość podstawowa czaszki MOR 1110 to 77,6 cm - mniejszy MOR 2951 ma 58,6 cm (Scannella i in., 2014) a większe okazy o dł. podstawowej czaszki ok. 90 cm są już wyraźnie dojrzalsze (USNM 1205, ok. 89 cm [Anderson, 1999] - Longrich i Field, 2012; YPM 1822, 88-93 cm [Forster, 1996b; Borkovic, 2013] - Longrich i Field, 2012; ROM 55380, ok. 90 cm [Farke, online 2012c; contra Borkovic, 2013 – zob. Farke, online 2012d] ma zwrócone ku przodowi rogi nadoczne). Szczególne wydłużenie rogów w tym stadium może wynikać z zaprzestania ich rozwoju a nawet ich resorpcji u starszych okazów (zob. Zmniejszanie ozdób w ontogenezie dinozaurów: Ceratopsidae). MOR 2597 - okaz o nieokreślonej przynależności taksonomicznej (zawierający jednak kość ciemieniową, więc prawdopodobnie diagnostyczny – Goodwin i Horner, 2010), pochodzący z górnych warstw formacji Hell Creek, ma rogi długości ok. 53,5 przy długości podstawowej czaszki 91,8 cm (stosunek długości: 0,58) (Scannella i in., 2014). Także holotyp T. prorsus, bardzo zbliżony w proporcjach do MOR 2597, wyróżnia się dość nietypową (tym razem znaczną), jak na swój gatunek, długością rogów.
(13) [4] zamknięte ciemiączka ciemieniowo-czołowe u dorosłych okazów.
(14) [24] bardziej wypukła krawędź kości łuskowej.
Komentarz
Większość cech (1-3, 5-14) zostało omówionych dość skrótowo we właściwym teście publikacji Scannelli i in. (2014) i suplemencie tekstowym, w tym omówieniu cech analizy kladystycznej. Bardziej szczegółowe omówienie cech oraz cechy 4 i 5 są pochodną analizy danych, zwłaszcza macierzy analizy kladystycznej i suplementu – zestawu danych z pomiarami.
Plezjomorficzny stan powyższych cech mają wcześniejsi przedstawiciele Triceratopsini. Dotyczy to cech 1, 2, 3, 6 i 8 u Eotriceratops (Wu i in., 2007; Scannella i in., 2014) i cech 6 i 8 materiału zaliczonego do Ojoceratops (Sullivan i Lucas, 2010). Podobna sytuacja występuje u Torosaurus latus i Titanoceratops. W przypadku zaś ?Torosaurus utahensis stwierdzić można jedynie, że kształt kości łuskowej (14) jest pośredni między Torosaurus latus a Triceratops (Maiorino i in., 2013b). Odmienny stan występuje u Regaliceratops (zaawansowany stan cech: 6, 12 i 14, bazalny u 10 i 13), który jest jednak wyspecjalizowaną, konwergentną z Centrosaurinae formą (Brown i Henderson, 2015) i według części analiz kladystycznych nie należy do Triceratopsini (Mallon i in., 2016; Dalman i in., 2021).
Część zmian jest peramorficznych (2?, 3?, 6, ?8, ?10, 13), natomiast inne – związane z potencjalnymi zmianami ontogenetycznymi w późnym stadium ontogenezy („Torosaurus”) mogą być pedomorficzne (1, 12 i 14).
MOR 3045 z górnych środkowych osadów jest w pewnych cechach (3, 4) bardziej zaawansowany niż inne z tego przedziału.
Dowody kladogenezy
Hipoteza o rozdzieleniu czasowym gatunków Triceratops może być uproszczeniem. Badania Scannelli i Fowlera (2009; 2014; Scannella i in., 2012; 2014) są ograniczone do niewielkiego obszaru okolic jeziora Fort Peck w Montanie, części formacji Hell Creek.
Lepszym przykładem prawdopodobnej kladogenezy jest UCMP 128561 (holotyp Ugrosaurus), który pochodzi z przedziału stratygraficznego T. ?sp. nov. (Scannella i Fowler, 2014; Scannella i in., 2014) a róg nosowy to niski guz, charakterystyczny dla wielu okazów z wcześniejszych osadów.
Hipoteza rozdzielenia czasowego nie pasuje odpowiada wielu innym danym stratygraficznym, w większości wątpliwych. Są to: (1) stratygrafia okazów z formacji Lance, zwłaszcza Hatchera i in. (1907), Lulla (1915) i Schlaikjera (1935), (2) prawdopodobna wysoka pozycja stratygraficzna BMRP 2006.4.1 mającego raczej plezjomorficzną budowę kości nosowej, (3) wyliczenia taksonów z dysertacji DePalmy (2010, s. 6-9), według którego w jednym stanowisku z górnej części formacji Hell Creek obecne są T. horridus i T. prorsus.
Interpretacja paleobiologiczna
Nie są znane przyczyny zmian ewolucyjnych w budowie Triceratops. Długość i masywność rogów i stowarzyszonych z nimi cech (2, 3, 6, 12) oraz połączeń między kością przedszczękową i nosową (4, 5) a być może nawet wszystkich cech tej części łba (1-11) może mieć związek ze sposobem, w jaki walczyły ze sobą Triceratops (por. Farke, 2004; Farke i in., 2009). Również zamknięcie ciemiączka (13) mogło mieć znaczenie biomechaniczne – wzmacnianie/usztywnianie czaszki (zob. Ciemiączka czołowo-ciemieniowe). Ewolucja budowy pyska mogą być również odzwierciedleniem zmian w odżywianiu (Longrich, online 2015). Longrich wskazuje jako analogię szybką ewolucję dziobów „zięb Darwina” z Galapagos.
Implikacje taksonomiczne
Podział linii anagenetycznej Triceratops na gatunki jest arbitralny. Scannella i in. (2014) określili okazy z górnych warstw jako T. prorsus a te z niższych do T. horridus, pozostawiwszy otwartą kwestię okazów pośrednich (ze górnych środkowych warstw). Te ostatnie w haśle określone jako T. ?sp. nov. (zob. niżej).
Diagnoza

Rodzaj Triceratops, jak i gatunek typowy (T. horridus), zdiagnozować należy następująco (cechy obecne u holotypu oznaczono gwiazdką - *):
1) Scannella i Horner, 2010 (zmodyfikowane z Ostrom i Wellnhofer, 1986 i Forster, 1996b) - pominięto cechy wynikające z uznania Torosaurus za synonim Triceratops oraz cechę T. prorsus a wiele z niżej wymienionych cech jest obecnych u znacznej części Ceratopsidae – diagnostyczne są cecha nr 5 i prawdopodobnie 6:
(1) Wydłużone rogi nadoczne (powstałe w wyniku połączenia kości zaoczodołowej i przedczołowej wcześnie w ontogenezie)* i (2) pojedynczy, zróżnicowanej budowy róg nosowy (tworzony przez kość nanosową - epinasale)*. (3) Róg nosowy łączy kompleks dziobowo-nosowo-przedszczękowy*. (4) Kształt ciemiączka ciemieniowo-czołowego (ang. frontoparietal fontanelle - Farke, 2010) jest zmienny, na ogół okrągły. (5) Kryza (złożona z k. ciemieniowej i łuskowej) w porównaniu do innych chasmozaurynów stosunkowo krótka, szeroka, wachlarzowata - przynajmniej u niezupełnie dorosłych okazów (zob. opis Torosaurus). (6) Mocno falisty środkowy grzbiet ciemieniowy staje się u dorosłych płaski. (7) Obecność epiparietales i episquamosales (rożków) na tylnych-bocznych krawędziach kryzy. (8) Kość najarzmowa (epijugale) na k. jarzmowej zrasta kompleks k. jarzmowej i kwadratowo-jarzmowej*.
2) Wu i in., 2007 - cechy odróżniające Triceratops od Eotriceratops (zob. opis):
Niski wyrostek trójkątny (sensu Forster, 1996a; Scannella i in., 2014, cecha 18) kości przedszczękowej - z górną krawędzią położoną znacznie pod wewnętrzną krawędzią okna międzyprzedszczękowego* oraz z zagłębieniem.
Pozostałe cechy podane przez Wu i in. (2007) są wątpliwe - zob. opis Eotriceratops.
3) Ott i Larson, 2010 - cecha odróżniająca Triceratops od Tatankaceratops (zob. opis):
Duże rozmiary: długość od dzioba do kłykcia potylicznego co najmniej ok. 90 cm u dorosłych okazów*.
4) Sullivan i Lucas, 2010 - cechy odróżniające Triceratops od Ojoceratops:
Kość łuskowa z dość ostro zakończonym tylnym końcem.
Co najmniej pozostałe cechy diagnostyczne Ojoceratops wydają się wchodzić w zakres zmienności Triceratops (Scannella i Horner, ryc. 3).
5) Longrich, 2011 oraz Longrich i Field, 2012 (podwójną gwiazdką - ** - oznaczono cechy podane tylko przez Longricha [2011]; część cech była wymieniana wcześniej w takim kontekście - np. Farke i Alley, 2006; Farke, 2011) - cechy kryzy odróżniające Triceratops od Torosaurus latus i ?Torosaurus utahensis (zob. opisy obu gatunków):
Kości ciemieniowe pozbawione okien. Tylna krawędź k. ciemieniowej gruba** i zadarta ku górze. Kość łuskowa krótka** i płaska (bez podłużnego wzniesienia wzdłuż łączenia z k. ciemieniową).
Obecność 5-7 epiparietales, w tym epiparietale położony centralnie, są prawdopodobnie obecne także u Torosaurus (zob. opis). Zob. też Torosaurus – diagnoza.
6) Brown i Henderson, 2015 - cechy odróżniające Triceratops od Regaliceratops:
Rogi nadoczne dłuższe od nosowego*. Episquamosales i epiparietales tych samych rozmiarów.
Ważność
Jak wynika z powyższej diagnozy, holotyp Triceratops horridus jest nieodróżnialny od Ojoceratops oraz Torosaurus.
Zachowane elementy typowych okazów Triceratops i Ojoceratops pokrywają się w niewielkim stopniu (przednia część kości łuskowych), co utrudnia lub uniemożliwia porównanie.
Od typowego okazu Torosaurus latus (YPM 1830) holotyp Triceratops horridus (YPM 1820) różni się otwartym ciemiączkiem ciemieniowo-czołowym (frontoparietal fontanelle - Farke, 2010) (cecha 13 w diagnozie T. horridus). Zamknięcie ciemiączka następuje w ontogenezie T. prorsus i Torosaurus latus, lecz prawdopodobnie nie T. horridus, gdyż nawet u całkowicie dojrzałych według Longricha i Fielda okazów USNM 2100 i USNM 2412 (holotyp Nedoceratops) ciemiączka są otwarte (Forster, 1996b). YPM 1820 nie ma jednej cechy najbardziej dojrzałych okazów według Longricha i Fielda (2012). Zamknięte ciemiączko ciemieniowo-czołowe mają również osobniki Torosaurus: znajdujące się w tym samym stadium rozwoju co YPM 1820 (MOR 1122 - Farke, 2006) i nieznacznie młodsze (ANSP 15192 - Farke, 2006).
Potencjalne synonimy
Na podstawie ww. cech diagnostycznych Triceratops prawdopodobnie nieodróżnialne są Nedoceratops hatcheri (= T. horridus), Ugrosaurus olsoni (= T. horridus), Tatankaceratops sacrisonorum (?= T. prorsus) i Ojoceratops fowleri (?= T. horridus) (zob. opisy i wpis o Nedoceratops poniżej). Nie jest pewne, na ile cechy podane jako diagnostyczne dla tych gatunków są rzeczywiście takimi (np. Longrich, 2011). Według niepublikowanych badań Ojoceratops i ?Torosaurus utahensis mogą być wcześniejszymi przedstawicielami linii anagenetycznej Triceratops (Fowler, 2010; Fowler i in., 2011). Torosaurus latus wpasowuje się w diagnozę Triceratops według Scannelli i Hornera (2010), ale nie Longricha (2011) oraz Longricha i Fielda (2012) a opisane w literaturze osobniki wykazują plezjomorficzną morfologię pyska T. horridus (holotyp, ANSP 15192, MOR 981, MOR 1122, MOR 3081).
Osobliwe okazy
Istnieje kilka pojedynczych okazów, które w różny sposób wykraczają poza standardową zmienność Triceratops. Poza ww., są to zwłaszcza AMNH 5116 i MOR 1120 (zob. Torosaurus: Hipotezy mieszane), MCZ 1102 (zob. T. eurycephalus), YPM 1822 (zob. wpis o holotypie T. prorsus) a także kontrowersyjny holotyp Tatankaceratops (zob. opis) oraz zbliżone do niego UCMP 136092 (bardzo krótkie rogi nadoczne i krótka, zwrócona silnie ku górze kryza, której dolna krawędź znajduje się nad linią zębów [Goodwin i Horner, 2010, ryc. 36.11]) i MOR 2972 (jeszcze krótsze rogi nadoczne) a także tzw. duranteceratops (omówiony niżej).
Trudno stwierdzić, czy cechy duranteceratops i innych osobliwych okazów uzasadniają wydzielanie taksonów. Materiał kopalny jest bogaty, zatem rośnie prawdopodobieństwo znalezienia osobników patologicznych.
Nedoceratops


Nedoceratops oparty jest na USNM 2412 - dość kompletnej czaszce.
Nedoceratops hatcheri w opinii niektórych naukowców jest ważnym gatunkiem (Forster, 1996b; Farke, 2011; Maiorino i in., 2013b). Według kolejnych jest młodszym synonimem Triceratops horridus (Longrich, 2010, Longrich i Field, 2012). Jeszcze inni uznają go za formę przejściową między młodszymi a w pełni wyrośniętymi Triceratops (tj. o budowie Torosaurus) (Scannella i Horner, 2010; 2011).
Nedoceratops ma mieszankę cech Torosaurus (np. budowa ciemiączek ciemieniowo-czołowych - ang. frontoparietal fontanelle) i Triceratops (np. płaska górna powierzchnia kości łuskowej) oraz pewne cechy pośrednie (np. długość kryzy), co skłania do przypuszczenia, że reprezentuje pośrednią formę rozwoju osobniczego - zob. Torosaurus: Okna i Formy pośrednie. Przeczy temu jednak inny (nieregularny i wydłużony) kształt okna.
Nedoceratops został zdiagnozowany następująco (Farke, 2011) (w nawiasach skróty nazw anatomicznych widoczne na rysunkach):
A. Autapomorfie:
1) róg nosowy (nh) niemal zupełnie nieodróżnialny od kości nosowej.
Komentarz: bardzo niskie, lecz nie aż tak jak u USNM 2412, rogi nosowe lub guzy obecne jest też u MOR 981, UCMP 128561 i USNM 4720 (Scannella i Horner, 2011). Powierzchnia ewentualnego rogu nie jest bardziej chropowata niż okolice (Farke, 2011, ryc. 1, 2, 4). Farke (2011) wskazuje, że róg jest obecny, o czym świadczy brak otwartego miejsca łączenia (szew) i oznak uszkodzeń, co kontrują Scannella i Horner (2011), stwierdzając, że gdy po utracie rogu połączenie kości zarastało, to szew nie będzie widoczny. Obecność rogu sugeruje kształt zachowanej kości - u niepołączonych z rogiem (epinasale) kościach nosowych Triceratops (np. Hatcher i in., 1907, ryc. 19; Mathews i in., 2009, ryc. 2; Schlaikjer, 1935, ryc. 3) widoczna jest wklęsła powierzchnia miejsca styku z rogiem, wyraźnie różniąca się od zaokrąglonego profilu kości USNM 2412. Możliwe, że róg został utracony we wczesnym etapie życia zwierzęcia, jak wskazywali Scannella i Horner (2011) czy Forster (1996a; 1996b). Horner i Goodwin (2008) zasugerowali, że utrata rogu i następujący po niej proces zdrowienia i remodelowania kości mogłyby prowadzić do powstania nieregularnego guza, widocznego u MOR 1122 i UCMP 128561. Te dwa okazy różnią się od USNM 2412 wyraźnie grubszą warstwą kości w miejscu rogu/guza w porównaniu do cienkiej kości USNM 2412 (Farke, 2006; Scannella i Horner, 2011, ryc. 2).
2) większa część rogów nadocznych (poh) zwrócona do góry (pod kątem prostym do linii zębów).
Komentarz: cecha widoczna u Triceratops YPM 1821 i okazu 1 z SMF (zob. ilustracje w haśle). U CNM 8862 taka budowa wynika zapewne z deformacji pośmiertnej (Forster, 1996b), czego autorka nie stwierdziła w zbadanym przez siebie holotypie Nedoceratops. Longrich i Field (2012) uznali natomiast, że po obu stronach czaszki Nedoceratops rogi nie są zwrócone ku górze, co wskazuje na uszkodzenie. Jest to widoczne na ryc. 1 w Farke (2011) – prawy jest bardziej pionowy, choć różnica w kącie ustawienia rogów jest minimalna. Na ryc. 2 jest zaś widoczne, że lewy róg jest skierowany bardziej na bok niż prawy. Zatem faktycznie występuje różnica w ich ustawieniu, co sugeruje deformację. Różny kąt nachylenia rogów zaobserwowano u innych ceratopsów (Lehman, 1990). Scannella i Horner (2011) sugerują, że w przypadku znacznej reorientacji rogów w ontogenezie, należy spodziewać się zróżnicowania w ich zwróceniu.
3) bardzo małe okna (pf) w kości ciemieniowej (p).
Komentarz: mogą być to patologie, często spotykane w kryzach ceratopsydów (Tanke i Farke, 2006). Świadczy o tym nieregularny kształt (Longrich i Field, 2012). Choć nieregularność otworu i otaczającej go cienkiej kości (Scannella i Horner, 2011) może nie wynikać z patologii a być rzeczywistą cechą anatomiczną, świadczy niewątpliwie, że to nie otwierające się okrągłe okno Torosaurus. Natomiast Farke (2011) uważa, że wydłużenie i znajdowanie się otworu w tylnej części kryzy odpowiada budowie innych chasmozaurynów z małymi oknami - Anchiceratops i Arrhinoceratops. Okna obu są jednak bardziej regularne – „wielokąty” tworzone przez nie mają wszystkie kąty wypukłe (Mallon i in., 2011; 2014), zaś Nedoceratops ma niektóre wklęsłe i jego okno jest bardziej wydłużone niż u ww. rodzajów. Farke (2011) wskazuje też, że tekstura kości wokół okna jest normalna, zatem nie powinna być to patologia. Obszar wokół otworu jest dość cienki – 0,8-1 cm (Farke, 2011). Lull (1905, s. 421) podał, że Gilmore, który preparował okaz, “nie jest wcale pewny” okna ciemieniowego.
B. Różnice w porównaniu do Triceratops:
4) brzuszny koniec kości łuskowej (sq) położony znacznie nad linią zębów.
Komentarz: obecne też u wielu Triceratops: AMNH 5116 (Matthew, 1915, ryc. 38), DMNH 48617 (Glut, 2006, s. 531), LACM 59049 (DeLuca, online 2011), MOR 3027 (Scannella i in., 2014, ryc. S1A), UCMP 113697 (Scannella i in., 2014, ryc. 1I), USNM 1201 (Hatcher i in., 1907, ryc. XLIII; Scannella i Horner, 2011, ryc. 5A).
C. Różnice w porównaniu do Torosaurus latus:
5) kształt kości łuskowej (sq) - szczególnie a) zredukowane wcięcie jarzmowe (jn) i b) brak zgrubienia przyśrodkowej krawędzi.
6) mała liczba różków (5?) na kości łuskowej (episquamosales).
Komentarz: rzeczywista różnica.
Istnieją też dwie kolejne cechy mogące być diagnostyczne:
7) Farke (2011) nie zawarł w diagnozie Nedoceratops obecności dodatkowych otworów (anterior temporal foramina) przy tylnym końcu kanałów grzbietowo-skroniowych (dorsotemporal channels – Farke, 2010), na które zwracała uwagę np. Forster (1996b), choć zaznaczył, że to cecha unikalna dla niego i niektórych Torosaurus latus i ?Torosaurus utahensis. Taki stan jest obserwowany tylko u części okazów Torosaurus latus (YPM 1831, lecz nie MOR 1122 – Farke, 2006), co sugeruje, że nie jest to cecha diagnostyczna – chyba że okazy należą do innych gatunków. Farke (2010, s. 1490) wskazuje, że z powodu znacznej rekonstrukcji tego obszaru u holotypu Nedoceratops oraz YPM 1831 rzeczywista budowa nie jest pewna (wcześniej [2006, s. 249] uważał, że rekonstrukcja wygląda na prawidłową). Jako unikalną cechę Farke (2010, s. 1490) podał zakończenie kanałów bliżej środkowej niż przedniej części okien grzbietowo-skroniowych (dorsotemporal fenestrae), choć na ryc. 5 w tej pracy nie widać, by kanały były wyraźnie przesunięte. Nie wydaje się to więc cechą diagnostyczną.
8) Okien obecnych w kościach łuskowych z reguły nie uznaje się za taksonomicznie istotne (np. Lull, 1933, s. 127; Farke, 2011). Choć jedno (lewe) z okien jest otoczone patologiczną tkanką (do 6,7 cm grubości), drugie (prawe) jest nie wykazuje tej cechy. Sugeruje to, że te prawe to dodatkowe okno o nieznanym pochodzeniu, obecne u wielu okazów różnych ceratopsydów (Tanke i Farke, 2006). Obecność okien nie wydaje się skorelowana z istnieniem charakterystycznego dla Torosaurus latus wgłębienia środkowej części kości łuskowej, w miejscu którego czasem pojawia się okno (Farke, 2006; online 2012a), gdyż kości łuskowe Nedoceratops wyglądają na płaskie (Farke, 2011, ryc. 2). Ponieważ żaden okaz Triceratops nie ma niepatologicznych otworów, sama skłonność do występowania dodatkowych okien może być jednak taksonomicznie istotna.
Podsumowując, poza cechami mogącymi wynikać z uszkodzenia (cecha 2) i patologii (cecha 3), niepewną budową przy tylnym końcu kanałów grzbietowo-skroniowych oraz oknami w kryzie o niepewnej diagnostyczności, wszystkie z cech mających różnić Nedoceratops i Triceratops są obecne także u niektórych okazów tego drugiego. Okazy te nie mają jednak pozostałych cech Nedoceratops albo są niewystarczająco kompletne. Holotyp Nedoceratops zdaje się mieć cechy T. horridus: 1) S-kształtny pysk, 2) nieco dłuższe niż T. prorsus rogi nadoczne, 3) otwarte ciemiączka ciemieniowo-czołowe i 4) prawdopodobnie niski (lub brakujący) róg nosowy (cechy 2 i 3 są obecne również u T. ?sp. nov.). Wskazuje to, że ten okaz jest przedstawicielem T. horridus. Jeśli Torosaurus i Triceratops to to samo zwierzę, to kolejne cechy diagnozy stają się nieistotne (np. zgrubienie przyśrodkowej krawędzi kości łuskowej - cecha 5a).
Geometryczne analizy morfometryczne Maiorino i in. (2013b) wykazały, że Nedoceratops mieści się w zakresie budowy (litera "a" oznacza wynik bez powiązaniu z rozmiarami a litera "b" w powiązaniu): 1) cała czaszka: 1a) poza Triceratops i Torosaurus, bliżej Triceratops, 1b) Torosaurus, 2) przód czaszki: 2a) poza Triceratops i Torosaurus, bliżej Triceratops, 2b) Torosaurus, 3) kryza: 3a) Triceratops, 3b) na styku Triceratops i Torosaurus, 4) kość łuskowa: 4a) Triceratops, 4b) Triceratops. Nedoceratops łączy zatem proporcje typowe dla obu rodzajów - niektóre zbliżają go do jednego a inne do drugiego. Interpretacja takich wyników może być rozmaita. Autorzy publikacji uważają, że świadczą one o odrębności taksonu. Alternatywnie można stwierdzić, że uwidaczniają one, że holotyp Nedoceratops stanowi formę przejściową między Triceratops a Torosaurus: ontogenetycznie (Scannella i Horner, 2010; 2011) - zob. opis Torosaurus lub ewolucyjnie (zob. Torosaurus: Trendy ewolucyjne Triceratops u Torosaurus). Wyniki nie są jednoznaczne, wykazują jednak znaczne zróżnicowanie Triceratops, od którego Nedoceratops nie jest bardzo oddalony. Liczne niewątpliwe okazy Triceratops również znajdują się w pewnym oddaleniu od pozostałych w różnych analizach - zwłaszcza AMNH 5116, MNHN 1912.20, MOR 1120 (T. horridus), MOR 1110 i YPM 1822 (T. prorsus). Wydaje się więc, że analizy Maiorino i in. (2013b) nie stanowią przeszkody do zaliczenia do Triceratops.
Jak udowadnia Farke (2011), Nedoceratops wykracza poza zmienność obserwowaną u Triceratops. Jednak to samo należałoby stwierdzić o wielu innych okazach przypisywanych Triceratops (zob. wyżej - #Osobliwe okazy).
Znany jest tylko jeden okaz o kombinacji nietypowych cech Nedoceratops – jego holotyp. Jak podaje nieoficjalnie Scannella (zob. Switek, online 2011), istnieje wiele nieopisanych osobników Triceratops, które mają cechy Nedoceratops.
Duranteceratops
Duranteceratops to nieformalna, nie podlegająca ICZN (art. 1.3.5) nazwa czaszki ceratopsyda. Nazwa ukazała się w magazynie Prehistoric Times w 2017 r. (nr 121, s. 7), w cudzysłowie (jako “Duranteceratops”) w podpisie fotografii rzeźby Keitha Strassera w dziale z listami czytelników (anonim, 2017; fotografia tej strony widoczna w Lampkins, online 2017). Mogła zostać nadana za sugestią Mendozy, odnoszącego się do dużego rogu nosowego i nosa Jimmy'ego Durante (Larson Paleontology Unlimited, online 2014a).

Okaz pochodzi z formacji Hell Creek - okolic Buffalo hrabstwa Harding w Dakocie Południowej (Orlowski, online 2012; Detrich, online 2013 – to ten sam okaz, co w później podawanych źródłach: por. np. z Johnson, T., online 2014c). Jest to dość kompletna lewa strona czaszki o długości ok. 223-225 cm i prawy róg nadoczny (N. Larson, online 2015b; online 2015c; online 2015d). Okaz został sprzedany (Detrich, online 2022). Detrich (w online 2015b) podaje, że okaz nie jest unikalny, tj. „szczątki tego tego rodzaju bestii pojawiały się w hrabstwie Harding od dekad, ale nie rozpoznano ich jako odrębnych od T. prorsus i T. horridus”. Okaz został częściowo zrekonstruowany: dodano u podstawy rogów nadocznych 2,5-5 cm, żuchwę (prócz kości przedzębowej – widoczna w Detrich, online 2021) i zęby (N. Larson, online 2015c), zrekonstruowane są fragmenty twarzoczaszki, w tym co najmniej obszar poniżej rogu nosowego i dziób, część kości szczękowej zawierająca zęby i część kryzy („Restoration consisted of a few small spots on the frill and face, a seam below the left brow horn, some work on the rostrum below the nasal horn and beak”) (Heritage Auctions, online 2018). Wklęsłość na wyrostku trójkątnym kości przedszczękowej (o ile nie jest to rekonstrukcja, choć wyrostek wydaje się obecny - Larson Paleontology Unlimited, online 2014a; online 2014b), obecna u Triceratops (w tym Nedoceratops [Farke, 2011]]) i Torosaurus, wskazuje, że okaz jest przedstawicielem Triceratopsini bardziej zaawansowanym niż Eotriceratops (Scannella i in., 2014), Titanoceratops i Regaliceratops (Brown i Henderson, 2015; ten ostatni według części analiz kladystycznych nie należy do Triceratopsini [Mallon i in., 2016]). Kość rostralna (dziobowa), o ile jest rzeczywista, jest bardzo mała, podobnie jak u T. horridus DMNH 48617. Być może jest ona u obu okazów niekompletna. Nietypową cechą jest m.in. niezrekonstruowany róg nosowy mierzący ok. 43 cm (N. Larson, online 2015b; pominięto wymiary podawane przed preparacją, tj. sprzed 18 lipca 2015 roku [N. Larson, online 2015a]), przy czym jest to wartość mierzona od górnej krawędzi otworu nosowego – rzeczywista długość rogu mogła wynosić ok. 38 cm. Róg jest absolutnie i prawdopodobnie proporcjonalnie największy spośród Triceratopsini - mierzy ok. 17% długości czaszki, co może być nieznacznie zawyżone, gdyż czaszka jest niekompletna. U Triceratops ww. wymiary to maksymalnie 15% i 36,6 cm, przy długościach czaszki zbliżonych do duranteceratops: MWC 7584 – róg 36,6 cm, czaszka 250 cm (Scannella i Horner, 2010; Scannella i in., 2014), MOR 2923 – róg 29 cm, czaszka ok. 200 cm (Horner i Lamm, 2011; Scannella i in., 2014). Róg nie jest jednak bardzo wydłużony, bowiem stosunek wysokości (ok. 38 cm) do długości (30,5 cm – N. Larson, online 2015b) to 1,25. Nadzwyczajna długość rogu jest cechą zbliżającą do Tatankaceratops (zob. Anageneza i heterochronia). Fowler (w Lampkins, 2017) sugeruje, że rozmiary rogu mogą wynikać z jego zgniecenia i rozciągnięcia – co wydaje się jednak mało prawdopodobne. Fowler (online 2015) uważa, że to prawdopodobnie bardzo stary T. prorsus, których – jak twierdzi, nie podając uzasadnienia poglądu – jest niewiele. Sugeruje również (w Lampkins, online 2017), że możliwe jest, że u rzeczywiście starych okazów róg mógłby być bardzo duży. Fowler wskazuje też (online 2015), że w całkowitym widoku bocznym róg nosowy nie jest aż tak duży i nie różni się on od innych T. prorsus. Rzadko spotykaną cechą jest brak zakrzywienia rogu, podobnie zbudowane są CCM 49-1, MOR 2923 i zwłaszcza BSM 1958G00003.00001. Wyrostek nosowy kości przedszczękowej wydaje się bardzo masywny i skierowany niemal pionowo. Może to być rezultatem nieprawidłowej rekonstrukcji, skoro „jakąś pracę” wykonano poniżej rogu nosowego („Restoration consisted of … some work on the rostrum below the nasal horn and beak”), obszar ten wydaje się zachowany jedynie częściowo (zob. Larson Paleontology Unlimited, online 2014a). Jak u T. prorsus, dolna krawędź górnej części kości nosowej jest prosta. Generalnie zatem długość rogu i budowa pobliskich kości jest podobna do T. prorsus, lecz cechy te są bardziej rozwinięte. Nozdrza zewnętrzne i przednia część czaszki są bardzo wysokie - bardziej niż u nieco go przypominających LACM 59049, YPM 1821, EM P15.1, BSM 1958G00003.00001 i LACM 151459, lecz wydaje się prawdopodobne, że jest to w pewnym stopniu wynik zgniecenia okazu. Rogi nadoczne mają ok. 66 cm (N. Larson, online 2015d), zatem są znacznie dłuższe niż róg nosowy, absolutnie trochę dłuższe niż u wszystkich T. prorsus i proporcjonalnie dłuższe niż u większości z nich - stosunek długości rogów do czaszki (29%) jest podobny do MOR 1604 (30%) (Scannella i in., 2014) i niższy niż u YPM 1822 (36%) (wymiary za Forster, 1996b; Hatcher i in., 1907; Borkovic, 2013). Na zdjęciu z preparacji koniec rogu nadocznego znajduje się nad nozdrzem zewnętrznym (Larson Paleontology Unlimited, online 2014b). Ponieważ podstawy rogów zostały częściowo zrekonstruowane (N. Larson, online 2015c), możliwe jest, że rogi zostały przesunięte zbyt ku tyłowi a powinny być zwrócone bardziej do przodu i nieznacznie krótsze. Budowa kryzy wydaje się charakterystyczna dla Triceratops (krótka, z zaokrąglonym brzegiem i brakiem zgrubienia), choć mogła ona zostać zgnieciona (Fowler oraz Mallon w Lampkins, online 2017) i/lub nieprawidłowo sklejona. To co widoczne jest jako boczna krawędź kryzy - kość łuskowa, może być równie dobrze tylną krawędzią - kością ciemieniową (por. zdjęcia z preparacji - Larson Paleontology Unlimited, online 2014a; online 2014b). Kryza została częściowo zrekonstruowana (Heritage Auctions, online 2018) i może być nadal niekompletna, w tym w przeniodolnej części, z nietypowym wyrostkiem. Jedyną cechą zbliżającą do Torosaurus jest prawdopodobne okno w kryzie (N. Larson, online 2015b; N. Larson w Biles, online 2015) - zob. Torosaurus: Trendy ewolucyjne Triceratops u Torosaurus. Okno jest nietypowo położone i nie wiadomo, czy jest rzeczywiste, czy to wynik niekompletności okazu. Cechą diagnostyczną ma być szeroki i płaski rowek na spodzie kości przedzębowej (Detrich, online 2015b). Wymagałoby to zbadania i porównania z innymi okazami. Doniesiono o podobnej kości, jednak jej pochodzenie nie zostało ujawnione (Maltese, online 2021a). Detrich (online 2017) opublikował zestawienie omawianego okazu wraz z zrekonstruowanym na jego wzór holotypem Tatankaceratops. Ze względu na ponad dwukrotną różnicę wielkości obu okazów ich przynależność do jednego gatunku jest mało prawdopodobna. Tatankaceratops nie jest młody, lecz maksymalnie dojrzały według analizy ontogenezy (Longrich i Field, 2012) mimo że czaszka ma ok. 1 m. Wydaje się, że mimo nieznacznie większego rogu nosowego niż u innych T. prorsus, okaz uznać należy – tak jak uważa Fowler – za starego (i dużego) osobnika tego gatunku. Ilustracje okazu dostępne są w następujących źródłach: Detrich, online 2013; Larson Paleontology Unlimited, online 2014a; online 2014b; Detrich, online 2015a, N. Larson, online 2015a; online 2015b; Detrich, online 2015c; O'Connell, online 2015; Clark, online 2015; Larson Paleontology Unlimited, online 2015a; online 2015b; "Blomman87", online 2017; Heritage Auctions, online 2018; Maltese, online 2021b; Detrich, online 2021.
Materiał kopalny ogólnie


Do rodzaju Triceratops zaliczyć można głównie okazy, które mają zachowaną kryzę (Farke, 2006; Longrich i Field, 2012). Geometryczne badania morfometryczne sugerują, że możliwe jest odróżnienie Triceratops, Nedoceratops i Torosaurus na podstawie kształtu przedniej (tj. bez kryzy) części czaszki (Maiorino i in., 2013b). Z kolei osobniki Triceratops można przypisać do gatunku głównie na podstawie morfologii pyska i rogu nosowego (Forster, 1996b; Longrich i Field, 2012; Scannella i in., 2014) – zob. diagnozę T. prorsus.
Ustanowienie Tatankaceratops (Ott i Larson, 2010) komplikuje przypisywanie okazów do Triceratops i jego gatunków (zob. opis). W zestawieniu poniżej uwzględniono okazy, które potencjalnie mogą być przedstawicielami Tatankaceratops (zawierające kryzę, ale nie rogi), gdyż rodzaj ten może być oparty na szczątkach osobnika z zaburzeniem rozwojowym.
Poza niżej wymienionym materiałem, istnieje wiele mniej lub bardziej kompletnych czaszek nie zaklasyfikowanych do gatunków. Niewątpliwy (stowarzyszony z diagnostycznymi czaszkami) oraz potencjalny (nie stowarzyszony z czaszkami) materiał pozaczaszkowy Triceratops jest liczny. Z reguły kręgi i kości kończyn Ceratopsidae nie są opisywane, lecz mogą być bardziej diagnostyczne, niż się powszechnie sądzi (Maidment i Barrett, 2011).
Już w 1906 r. Gilmore (s. 433) podał, że w północno-wschodniej części hrabstwa Converse (obecnie Niobrara) Hatcher z ekipą odkrył ponad 40 osobników zaliczanych do Ceratopsia. Zapewne zdecydowana większość z tych znalezisk, należały do Triceratops. Brown (1917, s. 281-282) twierdził, że podczas prac terenowych w Hell Creek w Montanie w latach 1902-1909 zidentyfikował "nie mniej niż 500 fragmentarycznych czaszek i niezliczone kości" należących do Triceratops, zaś Torosaurus był rzadki. Goodwin i Horner (2010, s. 555) na podstawie własnych doświadczeń polowych utrzymują, że nie jest to przesadzona liczba. Erickson (1966, s. 11) twierdził, że wszystkie okazy Triceratops - tj. ok. 200 - zaobserwowane w Hell Creek należą do T. prorsus. Zapewne większość ze wspominanego materiału nie została wydobyta a część może reprezentować potencjalne synonimy Triceratops. Dodson i in. (1990) wspominają o ok. 50 (zebranych) czaszkach. Horner i in. (2011) podają liczbę 72 osobników odkrytych w latach 1999-2009 w osadach formacji Hell Creek w północno-wschodniej Montanie. Scannella i in. (2011) wspominają o ponad stu czaszkach Triceratops z Montany; autorzy ci wliczają w to Torosaurus oraz okazy niediagnostyczne, gdyż uważają, że występował tam tylko jeden rodzaj chasmozauryna.
Legenda
Wyliczenie materiału kopalnego poniżej podzielono na pięć grup - okazów przypisanych na różnych podstawach: (1) budowy i wieku geologicznego, w tym okazy nieprzypisane formalnie do gatunku w literaturze, (2) budowy - przypisane w literaturze, (3) budowy, nieprzypisane w literaturze, (4) wieku geologicznego, (5) budowy, wbrew wiekowi geologicznemu. Jeśli nie wskazano inaczej, wzięto pod uwagę jedynie osobniki przypisane po 1996 r. (po publikacji Forster, 1996b) przez autorów wyróżniających dwa gatunki Triceratops (nie uwzględniono pracy Farke [1997], który użył pięciu cech diagnostycznych, z których pierwsza [długość rogów nadocznych] pokrywa się z propozycją Forster [1996b], druga [nachylenie rogów] zależna jest od ontogenezy [Horner i Goodwin, 2006], dwie kolejne [długość i wysokość kryzy] są wynikiem ontogenezy i różnic międzygatunkowych [Scannella i Horner, 2010; Maiorino i in., 2013b], natomiast rozstawienie rogów obecnie nie jest uznawane za cechę diagnostyczną; proponowano, że zależy od płci). Okazy mające diagnostyczną budowę zaawansowanych Triceratops (T. ?sp. nov. i T. prorsus), lecz o budowie niepozwalającej na zaliczenie go do rodzaju, przypisano do tych gatunków z oznaczeniem „$”, gdyż u Torosaurus nie stwierdzono obecności takich cech, zwłaszcza dużego rogu (jego obecność byłaby argumentem, że jest to synonim Triceratops, choć nie można też wykluczyć konwergencji). Okazy mogące należeć ewentualnie do Tatankaceratops oznaczono znakiem „^”, lecz nie wzięto takiej ewentualności przy zliczeniu okazów. Przyjęto, że przy porównaniu z Tatankaceratops, przypisanie do Triceratops jest niewątpliwe, jeśli okaz spełnia którąś z następujących przesłanek: (1) ma diagnostyczne cechy Triceratops horridus: 1-5 lub 9 (Tatankaceratops ma cechy zaawansowanych Triceratops, zatem plezjomorficzna budowa wyklucza zaliczenie okazu do Tatankaceratops), (2) jest duży - odległość od dzioba do kłykcia potylicznego powyżej 100 cm (różnica między rozmiarami największego [EM P15.1] i najmniejszego dorosłego Triceratops [YPM 1822] to 160%, co przy szacowanej tej długości holotypu Tatankaceratops, wynoszącej 64 cm, pozwala przypuszczać, że największe Tatankaceratops mogły mieć ten wymiar o wartości ok. 100 cm; w kwestii ww. wymiarów zob. Torosaurus: Największe ceratopsy), (3) róg nosowy niewydłużony, (4) róg nosowy krótszy niż 20% długości od dzioba do kłykcia potylicznego, (5) róg nosowy wydłużony, lecz dłuższy niż 23 cm (omówione w pkt 1 różnice wielkości pozwalają na przypuszczenie, że u największych Tatankaceratops róg mógł mieć 23 cm), (6) rogi nadoczne dłuższe niż 23 cm (jak w pkt 4), (6) rogi nadoczne proporcjonalnie znacznie dłuższe niż u holotypu Tatankaceratops (stosunek do długości od dzioba do kłykcia potylicznego powyżej 0,35 – przyjęto arbitralnie; u holotypu Tatankaceratops wynosi 0,23), (8) rogi nadoczne dłuższe niż u holotypu Tatankaceratops, lecz skierowane ku tyłowi, (9) rogi nadoczne zakodowane w Longrich i Field (2012, cecha 12) jako masywne, (10), pochodzi z dolnej połowy formacji Hell Creek (Tatankaceratops ma cechy zaawansowanych Triceratops, obecnych w górnej połowie formacji), (11) określony jako „duży”, (12) określony jako „dorosły”.
Okazy dokładniej opisane w literaturze pogrubiono a holotypy podkreślono. Okazy zilustrowane w informatywny sposób w literaturze lub internecie zaznaczono kursywą. Podano miejsce zilustrowania okazu. Gwiazdka oznacza zilustrowanie w niniejszym haśle, obok wpisu o okazie; podwójna gwiazdka - w innym miejscu hasła; "O" oznacza zilustrowanie w dużym rysunku w rozdziale #Ontogeneza. Wybrano ilustracje najlepszej jakości (ryciny z Hatcher i in. [1907] pochodzą czasem z wcześniejszych prac Marsha i często były reprodukowane w kolejnych pracach). Pojedynczy znak zapytania przed okazem wskazuje, że nie jest on zilustrowany lub z innych (w tym przypadku podanych) powodów nie można potwierdzić prawidłowości przypisania do rodzaju. Podwójny lub potrójny znak zapytania oznacza wątpliwości co do przypisania wyrażone w literaturze. Znak zapytania w nawiasie oznacza niepewność przypisania do gatunku. Zasady te nie dotyczą znaków zapytania u osobników nieprzypisanych w literaturze, gdzie podano powody dla wątpliwości przypisania. Dla tych okazów wzięto pod uwagę tylko te o niewątpliwej autentyczności oraz często z jakiś powodów warte uwagi i ze znanymi numerami katalogowymi. Podano także cechy diagnostyczne, które przesądziły o przypisaniu do gatunku (zob. diagnozy gatunków). Znaki zapytania po skrócie instytucji oznaczają nieznany numer katalogowy.
Lokalizacje ogólnie
Według danych przedstawionych niżej (w omówieniach gatunków) Triceratops znany jest z:
USA: formacja Lance (Wyoming), formacja Hell Creek (Montana, Dakota Południowa, Dakota Północna), formacje Denver i Laramie (Colorado); Kanada: formacja Frenchman (Saskatchewan) i Scollard (Alberta)
Niewątpliwe szczątki Triceratops znane są także z następujących osadów: formacja Evanston, Wyoming (Dodson i in., 2004; Goodwin i Horner, 2010), a wątpliwe (za Weishampel i in., 2004) także z: formacja Kaiparowits, Utah (cf. Triceratops sp.) (kampan); formacja Medicine Bow, Wyoming (=Chasmosaurinae indet.); formacja Ferris, Wyoming (=Chasmosaurinae indet.).
T. horridus
Diagnoza
Zob. diagnozę rodzaju.
Na podstawie Scannella i in. (2014) cechami pozwalającymi odróżnić T. horridus od pozostałych gatunków (zob. „dolne osady”) są (gwiazdką oznaczono cechy obecne u holotypu):
(1) długi pysk - stosunek długości od dzioba do podpórki nosowej kości przedszczękowej (narial strut) powyżej do długości od dzioba do kłykcia potylicznego powyżej 0,28.
(2) [3] pochylony ku tyłowi wyrostek nosowy kości przedszczękowej - kąt między nim a „rozporą nosową” wynosi poniżej 131 stopni.
(3) [14] wąski wyrostek nosowy kości przedszczękowej - stosunek wysokości (mierzonej od najniższego punktu artykulacji z kością nosową) do szerokości (długości) najwęższej części wynosi powyżej 1,57.
(2-3) obie te cechy, zwłaszcza ta pierwsza, powodowały, że pysk miał opisaną przez Forster (1996b, cecha 4) budowę, tj. „wydłużony, S-kształtny pysk”*.
(3) [21] wąska tylnobrzuszna powierzchnia dolnotylnego wyrostka kości przedszczękowej.
(5) [22] szeroka powierzchnię styku z kością nosową dolnotylnego wyrostka kości przedszczękowej.
(6) [7] krótki i niewydłużony róg nosowy - do 15,5 cm długości. Stopień wydłużenia rogu (długość/szerokość) do 1,5. Długość poniżej 0,15 długości od dzioba do kłykcia potylicznego (u okazów niebędących młodymi, ze słabo rozwiniętymi rogami).
(7-8) [13] guzek na tylnej powierzchni rogu nosowego i [10-11] guzek formowany przez tylną część rogu nosowego i przednią część kości nosowej. Obecne także u T. ?sp. nov..
(9) krótka kość nosowa – stosunek jej długości do długości od dzioba do kłykcia potylicznego u większych okazów (druga długość ok. 1 m) poniżej 0,38.
(10) [9] krótka i łukowata wygięta dolna krawędź górnej części kości nosowej*. Obecne także u T. ?sp. nov..
(11) [12] wyraźny przedniobrzuszny wyrostek kości nosowej.
(12) [1] rogi nadoczne dłuższe niż 0,45 długości od dzioba do kłykcia potylicznego - u większych okazów (o tej długości większej niż 50-90 cm). Obecne także u T. ?sp. nov..
(13) [4] otwarte ciemiączka ciemieniowo-czołowe (frontoparietal fontanelle - Farke, 2010) u dorosłych okazów (Forster, 1996b)*. Nieznane u T. ?sp. nov..
Materiał kopalny
Holotyp to YPM 1820 - niekompletna, mocno zwietrzała czaszka (brak zwłaszcza większości kryzy) o szacowanej długości ok. 2 m (Lull, 1933, s. 116) (Marsh [1889b] ocenił ją początkowo na co najmniej 2 m, „nie licząc rogowego dzioba” a później [1889c] na ponad 2,4 m [8 stóp]). Pochodzi od starego osobnika (np. Hatcher i in., 1907; Longrich i Field, 2012). Czaszka opisana w: Hatcher i in., 1907, s. 117 i n. Ilustracje: *; Tsuihiji, 2010; Farke, online 2018; Scannella, 2020, ryc. 8. Lokalizacja: wcześniejsze osady formacji Lance ("środkowa część górnej połowy Laramie"), hrabstwo Niobrara, Wyoming (Hatcher i in., 1907; Ostrom i Wellnhofer, 1986).
(1) Materiał przypisany na podstawie morfologii i wieku geologicznego (zob. #Legenda):


- MOR 1120 - niedorosły osobnik (subadult - Longrich i Field, 2012; Horner i Goodwin, 2006) z czaszką dł. całkowitej ok. 165 cm (Horner i Goodwin, 2006) i podstawowej 98 cm (Scannella i in., 2014). Ma niewielki róg nosowy, jak T. horridus - jest on mniej wydatny niż u mniejszych od MOR 1120 okazów z górnej części formacji Hell Creek (T. prorsus) - MOR 1110 i MOR 2951. Na fotografii w pracy Hornera i Goodwina (2006, ryc. 1d) oraz okaz Goodwina i Hornera (2010, ryc. 36.12) ma krótki dziób, jak T. prorsus, lecz na nowszej fotografii (Rogers i Rafferty, online 2010) oraz w pracy Scannelli i in. (2014) pysk jest długi i ma esowaty kształt, jak u T. horridus. Według Hornera i Goodwina (2006) okaz jest kompletny, lecz po publikacji tego badania został ponownie zrekonstruowany (Scannella, prywatna korespondencja VII.2014). Ilustracje: *; O; Horner i Goodwin, 2006, ryc. 1d; Scannella i in., 2014, ryc. 1J; Britannica, online 2010; Scannella, 2020, ryc. 5C; MOR online 2019b. Lokalizacja: górne dolne osady formacji Hell Creek, Montana (Horner i in., 2011; Scannella i Fowler, 2014);
- ? MOR 3011 - zawiera przynajmniej część kości ciemieniowej, róg nosowy (Scannella i Fowler, 2014), kość przedszczękową, nosową i niekompletną żuchwę (Scannella i in., 2014). Fragmenty kryzy są "dość grube" (Scannella i in., 2014), co sugeruje, że to Triceratops, lecz w analizach kladystycznych Scannelli i in. (2014) znajdował się w pobliżu Torosaurus, mimo że jest późniejszym od nich geologicznie okazem - wynika to jednak prawdopodobnie z jego niekompletności (w analizie kladystycznej Scannelli i in. nie zawarto cechy dotyczącej grubości kryzy). W każdym razie MOR 3011 wykazuje plezjomorficzną budowę pyska jak u T. horridus i Torosaurus (Scannella i in., 2014). Autorzy ci podają, że to okaz "bardzo duży młody lub dojrzewający" („either a very large juvenile or a subadult”). Oszacowana na podstawie długości kości zębowej długość od dzioba do kłykcia potylicznego to ok. 88 cm (Scannella i in., 2014). Ilustracje: Scannella i in., 2014, ryc. S2C. Lokalizacja: dolne środkowe osady formacji Hell Creek, hrabstwo Garfield, Montana (Scannella i Fowler, 2014);
- USNM 4720 - holotyp T. obtusus. Jedna z największych czaszek Triceratops (zob. Torosaurus: Największe ceratopsy), opisana w: Hatcher i in., 1907, s. 140 oraz Gilmore, 1920, s. 98 i n. oraz kręg (Gilmore, 1920. Ilustracje: *; Lull, 1933, ryc. 41; Glut, 1997, s. 924; Pence, online 2013; USNM, online. Lokalizacja: wcześniejsze osady formacji Lance ("środkowa część Laramie"), hrabstwo Niobrara, Wyoming (Hatcher i in. 1907; Ostrom i Wellnhofer, 1986).
(2) Materiał przypisany w literaturze na podstawie morfologii (zob. #Legenda):
- Forster, 1996b:
- AMNH 5116 – prawie kompletna czaszka, lecz kryza jest niekompletna (brak okien jest jednak widoczny) i nie zachował się dziób (Matthew, 1915, s. 110), którego rekonstrukcja jest zapewne błędna, gdyż nie odpowiada kształtowi dzioba innych osobników – jest za długi. Według Forster (1990) miał to być odrębny gatunek. Okaz jest zmontowany w szkielet z kilkoma innymi osobnikami (Osborn, 1933). Maiorino i in. (2013a) podali nr 5516, który zapewne jest pomyłką. Ilustracje: ** - Taksonomia; Osborn, 1933; Scannella i Horner, 2010, ryc. 5a. Lokalizacja: formacja Lance, hrabstwo Niobrara (lub Weston - Lull, 1933, s. 123), Wyoming (Forster, 1996b);
- FMNH P12003 - czaszka. Ilustracje: *; Lull, 1933, tabl. XIIIa; Glut, 1997, s. 31 i 930; Glut, 2008, s. 147 i s. 172, Ford, online - Triceratops. Lokalizacja: formacja Hell Creek, Chalk Buttes, poł.-zach. Montana (Lull, 1933; Forster, 1996b);
- SDSM 2760 – zapewne jest to ten samo okaz, co SDSM P271, opisany i przypisany do T. horridus przez Lulla (1933, s. 117). Oba są bardzo podobne, choć pochodzić mają z różnych lokalizacji – wg Forster (1996b) to zachodnia Montana, lecz pozostałe źródła podają Dakotę Południową. Numer P271 widnieje tylko w publikacji Lulla (1933), w innych pracach podaje się 2760 (m.in. Forster, 1996b; Lehman, 1998; Farke, 2006). Ilustracje: *; Lull, 1933, tabl. XIIIb – jako P271; Forster, 1996b, ryc. 9a; Ford, online - Triceratops. Lokalizacja: Short Pine Hills – okolica Camp Crook, hrabstwo Harding, Dakota Południowa (Lull, 1933; SDSM, online 2017);
- USNM 1201 - holotyp T. elatus. Czaszka opisana w: Hatcher i in., 1907, s. 135-136. Ilustracje: *. Lokalizacja: formacja Lance, Lance Creek, hrabstwo Niobrara, Wyoming (Ostrom i Wellnhofer, 1986);
- USNM 2100 – czaszkę umieszczono wraz z okazem USNM 4842 w zmontowanym szkielecie (Ostrom i Wellnhofer, 1986, s. 130). Krótki róg nosowy wygląda na kompletny i wskazuje, że to T. horridus, jednak Gilmore (s. 434 oraz w Schuchert 1905, s. 459) podaje, że róg nie jest w części lub w całości zachowany. Okaz ma długie rogi nadoczne i otwarte ciemiączka czołowo-ciemieniowe mimo zaawansowanego wieku (Longrich i Field, 2012). Ilustracje: Schuchert, 1905 - czaszka; Gilmore, 1906 - czaszka; Hatcher i in., 1907, tabl. XXX; Swiderski, online 2008. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Ostrom i Wellnhofer, 1986);
- USNM 4928 - holotyp T. calicornis – dość kompletna czaszka i materiał pozaczaszkowy, m.in. 2 kręgi szyjne i 10 grzbietowych, kość krzyżowa i kości miednicy (Gilmore, 1920). Zachowała się początkowo połowa kości ciemieniowej, lecz obecnie istnieją jedynie jej fragmenty (Hatcher i in., 1907, s. 138-139). Kości łuskowe nie zdradzają obecności podłużnego zgrubienia, więc nie jest to Torosaurus. Okaz opisany w: Hatcher i in., 1907, s. 136 i in. oraz Gilmore, 1920, s. 101 i n. Ilustracje: *; Hatcher i in., 1907, tabl. XXXVIII; Forster, 1996b, ryc. 3a; Glut, 1997, s. 922. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Hatcher i in. 1907; Ostrom i Wellnhofer, 1986);
- Glut, 2006:
- DMNH 48617 - wypukła powierzchnia kości łuskowej wskazuje, że to Triceratops (Shay, online 2008). Ilustracje: Glut, 2006, s. 531; Carpenter, 2006, ryc. 15.5; Ford, online - Triceratops; National Science Foundation, online 2010. Lokalizacja: formacja Denver/Sequence D1, Colorado (Glut, 2006; Fowler, online 2017);
- TCM 2001.93.1 ("Kelsey") zawiera praktycznie kompletne: czaszkę długości ponad 2,11 m (BHI online b; ponad 2 m wg Otta i Larsona [2010, s. 211]; wg Moliny Péreza [online 2019] 2,26 m), ciąg kręgów i żebra. Brak większości ogona, kończyn przednich i stóp (BHI, online b). Szacowana długość ciała to 6,1 m (BHI, online b). Ilustracje: Glut, 2006, s. 114 i s. 530; Glut, 2008, s. 242, s. 635, s. 636; Ford, online - Triceratops; "Anselmo", online 2010; BHI, online b. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (P. Larson i in., 2004);
- Goussard, 2006:
- MNHN 1912.20 – czaszka ze znacznie zrekonstruowaną kryzą (Goussard, 2006; Johnson, online 2022), prawa kość łuskowa nie zdradza obecności diagnostycznego dla Torosaurus zgrubienia. Goussard (2006) zaliczył okaz do tego gatunku, gdyż uznał, że to jedyny gatunek. Długa, S-kształtna przednia część pyska i mały róg nosowy wskazują, że to w istocie T. horridus. Ten sam numer katalogowy ma izolowany róg (Goussard, 2006). Czaszka opisana w: Goussard, 2006. Ilustracje: Goussard, 2006; Johnson, online 2022. Lokalizacja: formacja Lance, Lance Creek, hrabstwo Converse (Niobrara?), Wyoming (Goussard, 2006);
- Sampson i in., 2010:
- USNM 1205 - według Longricha i Fielda (2012) T. sp.. Większość pyska i rogi nie zachowały się, lecz zagięta dolna krawędź górnej części kości nosowej (USNM, online 2014) sugeruje, że jest to T. horridus. Ilustracje: USNM, online 2014. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Ostrom i Wellnhofer, 1986; Anderson, 1999);
- Ott i Larson, 2010:
- ROM 55380 - czaszka niedorosłego osobnika (subadult) (Maiorino i in., 2013b). Róg nosowy wydaje się dość wydłużony, lecz został on zrekonstruowany (Woollatt, online 2016). Farke (online 2012d, s. 6 – identyfikacja okazu według instytucji i wymiarów podanych w Farke, online 2012c) podaje, że mózgoczaszka jest w rekonstrukcji przesunięta zbyt daleko do tyłu. Ilustracje: Woollatt, online 2016. Lokalizacja: formacja Hell Creek, hrabstwo Harding, Dakota Południowa (Maiorino i in., 2013b; BHI, online c);
- Longrich, 2010 / Longrich i Field, 2012:
- USNM 2412 - holotyp Nedoceratops hatcheri – zob. #Nedoceratops. To dość dobrze zachowana czaszka o długości 1,8 m, opisana w Farke, 2011. Ilustracje: *; ** - #Nedoceratops. Lokalizacja: formacja Lance, okolice Lightning Creek, hrabstwo Niobrara, Wyoming (Hatcher, 1905; Ostrom i Wellnhofer, 1986);
- Lyson i Longrich, 2011:
- ? UCMP 140416 – duży okaz (por. wymiary z Chinnery, 2004), zawierający kryzę, której kształt określono jako „siodłowaty” („fan-shaped” - Horner i Goodwin, 2006, tab. 2), co sugeruje prawdopodobnie Triceratops. Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (Lyson i Longrich, 2011; UCMP, online);
- Longrich i Field, 2012:
- YPM 1828 - holotyp T. "ingens" (nomen nudum) - dość kompletna czaszka o szacowanej długości ok. 240 (Longrich i Field, 2012) lub ponad 244 cm (Lull, 1933, s. 117 - wówczas największa czaszka Triceratops). Zawiera co najmniej rogi nadoczne i kości: nosową, przedczołową, czołową, łzową, łuskową z episquamosales, ciemieniową z epiparietales (Longrich i Field, 2012), jarzmową (Farke i in., 2009) oraz kręgi (Lull, 1933). Forster (1996b) uznała go za Triceratops a Longrich i Field (2012) za T. horridus. Raczej niewielki róg nosowy i stosunkowo silnie zakrzywiona dolna krawędź górnej części kości nosowej, widoczna na ryc. na s. 928 w książce Gluta (1997), sugeruje, że identyfikacja jako T. horridus jest prawidłowa – ile zakrzywienie nie wynika z kąta zrobienia zdjęcia i jest to faktycznie ten okaz – podpisano go bowiem „?Triceratops ingens, YPM 2157, holotype rostrum”. Ilustracje: ? Glut, 1997, s. 928; fragment kości łuskowej - Farke i in., 2009, ryc. 2B. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Ostrom i Wellnhofer, 1986);
- Maiorino i in., 2013a/2013b:
- ??? BHI 4772 ("Billy" - Kato, online 2019) - czaszka niedorosłego osobnika (subadult) o długości 2,3 m, zaliczona do cf. Torosaurus (Ott i Larson, 2010; taką klasyfikację podają też Evans i in. [2012]). Według Maiorino i in. (2013b) to Triceratops horridus, których analizy morfometryczne sugerują, że to prawdopodobnie rzeczywiście Triceratops, lecz zbliżony budową do Torosaurus (analizy czaszki i kryzy) lub forma pośrednia między nimi (przód czaszki) - w tym bliższa Triceratops (kość łuskowa). Odlewy bardzo podobnego okazu (por. np. gramunion.com, online; Larson, online 2015) są wystawione w kilku instytucjach: ANSP ([Rolfes, online 2017]), ASU ([ASU, online 2012]), Oxford University Museum of Natural History (zdjęcie obok), Vienna Natural History Museum ([Evenor, online 2014]) i Drexel University ([iknowdino.com, online 2015]). Ilustracje: BHI, online 2015; Davis, online 2012. Lokalizacja: formacja Hell Creek, hrabstwo Corson, Dakota Południowa (Maiorino i in., 2013b; Maiorino i in., 2017; BHI – za Kato, online 2019);
- ^ (?) DMNH 1018 - przynajmniej kość łuskowa, według badań morfometrycznych Triceratops sp. (Maiorino i in., 2013b). Lokalizacja: formacja Hell Creek, Dakota Południowa (Maiorino i in., 2013b);
- ^ (?) FMNH PR2414 - przynajmniej kość łuskowa, według badań morfometrycznych należąca do Triceratops (Maiorino i in., 2013b). Lokalizacja: formacja Hell Creek, Montana (Maiorino i in., 2013b);
- HMNS PV.1506 ("Lane"; wcześniej BHI 6273 [m.in. P. Larson i in., 2007; Hendrickx i Bell, 2021]; według strony internetowej BHI [online a] okaz ma numer 6220; taki też numer podają Maiorino i in. [2013b] oraz Ott i Larson [2010]; często podawany numer HMNS 2006.1743.00 jest prawdopodobnie tymczasowy ["GetAwayTrike", online 2021]; w każdym razie odcisk skóry ma numer PV.1506 [Bell i in., 2022; Hendrickx i Bell, 2021]) - niemal kompletny, nieopisany okaz, zachowany z wieloma odciskami skóry (Leap, online 2012a). Z elementów czaszki brakuje jedynie kości ciemieniowej (BHI, online a). Kształt rogu nosowego i pyska wskazuje, że to T. horridus. Okaz został odkryty niedaleko "mumii" Edmontosaurus annectens (AMNH 5060) (P. Larson i in., 2007). Ilustracje: *; BHI, online a; Kato, online 2021. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (P. Larson i in., 2007; Leap, online 2012b);
- D’Anastasio i in., 2022:
- "Big John" - niekompletny duży szkielet (jedyna przekraczająca 1 m długości kość łuskowa Triceratops – 1,12 m), złożony z ponad 200 identyfikowalnych elementów (D’Anastasio i in., 2022). Ilustracje: D’Anastasio i in., 2021, ryc. 1; D’Anastasio i in., 2022, ryc. 1. Lokalizacja: formacja Hell Creek, hrabstwo Perkins, Dakota Południowa (D’Anastasio i in., 2022).

(3) Materiał nieprzypisany w literaturze (zob. #Legenda).:
- ^ ? BCGM 10272 – co najmniej kość łuskowa i kłykieć potyliczny, zaliczone do T. horridus w niepublikowanym badaniu histologicznym (Brown i Smith, 2017). Autorzy ci zidentyfikowali okaz jako dorastający (ang. sub-adult) posiłkując się m.in. grubością kryzy, która według nich u dorosłych staje się cienka i uzyskuje okna (tj. uzyskuje budowę Torosaurus). Sugeruje to, że u BCGM 10272 kryza jest gruba, zatem należy do Triceratops. Lokalizacja: Dakota Północna (Melvin, online 2016);
- (?) BMRP 2006.4.1 ("Homer") - jeden z trzech okazów z "Homer site". Zawiera kość nosową i rogi nadoczne (Mathews i in., 2009) a według fotografii okazu również kość rostralną, lewą kość jarzmową, prawą szczękową, kości zębowe i kryzę (Hunt-Foster, online 2009; BMRP, online 2010; Johnson, online 2014d). Stanowisko Homer zawiera trzy młode, zidentyfikowane jako Triceratops na podstawie braku okien w znalezionych w stanowisku kościach ciemieniowych (Mathews i in., 2009). BMRP 2006.4.1 to prawdopodobnie T. horridus na podstawie zagiętej dolnej krawędzi górnej części kości nosowej (zob. Komentarz do cech w ewolucji Triceratops: cecha 10). Skoro przynajmniej jeden z okazów należy do T. horridus, to prawdopodobnie także kolejne dwa należą do tego samego gatunku. Ilustracje: BMRP, online 2010; Mathews i in., 2009, ryc. 2A, C i F; Hunt-Foster, online 2009; Farke, 2014, ryc. 2e; Johnson, online 2014d. Lokalizacja: górne środkowe(?)-górne(?) warstwy formacji Hell Creek (zob. #Dowody kladogenezy), hrabstwo Carter, Montana (Mathews i in., 2009);
- ? BMRP 2006.4.307 - drugi okaz z "Homer site", zawiera m.in. róg nadoczny. Ilustracje: Mathews i in., 2009, ryc.2B; zob. wyżej;
- ^ ? okaz z BMRP - trzeci okaz z "Homer site", zob. wyżej;
- (?) CCM 49-1 - czaszka. Zawiera kryzę i rogi nadoczne (Farke, 1997). Farke i in. (2009) podają, że okaz zawiera kości: nosową, jarzmową, ciemieniową i łuskową, podczas gdy drugi okaz z CCM wyliczony przez tych autorów zawiera tylko kości nosowe. Maiorino i in. (2013b) użyli tego okazu do badań morfometrycznych (więc jest on praktycznie kompletny i artykułowany) i zaliczyli go do dorosłego T. prorsus. Wszystkie te dane wskazują, że jest to zapewne okaz na zdjęciu obok. Ma długie rogi nadoczne, jak T. horridus i T. ?sp. nov.. Dolna krawędź dolnej części kości nosowej jest krótka i zakrzywiona, co wskazuje niewątpliwie na T. horridus, choć róg nosowy jest duży - co jest charakterystyczne dla zaawansowanych Triceratops - aczkolwiek nie wydłużony – wydaje się krótszy (mniej wydłużony) niż u T. ?sp. nov. i T. prorsus. Może być zaawansowanym T. horridus lub formą pośrednią między nim a T. ?sp. nov. (zob. Komentarz do cech w ewolucji Triceratops: cecha 6). Ilustracje: ?*; ?Johnson, online 2014a. Lokalizacja: formacja Hell Creek, Montana (Maiorino i in., 2013b);
- ? FPDM-V-9677 – opisana puszka mózgowa oraz "kość nosowa, przedszczękowa, szczękowa, "rostrum", zaoczodołowa, jarzmowa, kwadratowa, ciemieniowa i łuskowa" (Sakagami i Kawabe, 2020). Czaszka wygląda na dość kompletną, ale niektóre obszary - z pewnością przed i za oczodołem oraz róg nosowy wyglądają na zrekonstruowane. Budowa kryzy jest typowa dla Triceratops a kształt pyska sugeruje T. horridus. Ilustracje: Sakagami i Kawabe, 2020. Lokalizacja: formacja Hell Creek, Ziebach, Dakota Południowa (Sakagami i Kawabe, 2020);
- (?) okaz z GRM ("Grath") - czaszka zaliczona do T. horridus (GRM, online 2013). Praktycznie kompletna kryza z budową diagnostyczną dla Triceratops i przednia część kości nosowej sugerująca, że róg nosowy musiał być niewielki, wskazują na poprawność takiej identyfikacji. Ilustracje: GRM, online 2012; online 2013; Arnold, online 2014, Peterson, online. Lokalizacja: okolice rzeki Grand – ranczo Schmidtów, Dakota ?Południowa (GRM, online 2012);
- ? okaz z MOS ("Cliff") - dość kompletny szkielet (brak niektórych kręgów, żeber i kości kończyn) mierzący ok. 7 m (MOS, online). Czaszka została znacząco zrekonstruowana (Farke, online 2016). Okaz ma pochodzenie komercyjne i może zostać utracony przez MOS (Timmons, online 2014). Ilustracje: MOS, online; "Daderot", online 2010. Lokalizacja: formacja Hell Creek, Dakota Północna (Timmons, online 2014);
- (?) LACM 27428 – czaszka z kryzą pozbawioną okien, co potwierdza przynależność do Triceratops. Róg nosowy wydaje się mały jak na T. ?sp. nov. i T. prorsus a duży jak na T. horridus - najbardziej przypomina wydatne rogi AMNH 5116 i SDSM 2760, lecz jest mniejszy niż u CCM 49-1. Sampson i in. (2010) zaliczyli go do T. horridus. Ilustracje: Glut, 2006, s. 116; LACM, online. Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (LACM, online);
- okaz z LASM ("Jason") - czaszka o długości ok. 2,2 m, która jest niemal kompletna (LASM, online 2017; WBRZ, online) - inny kolor ma tylna prawa strona kryzy, co sugeruje, że ta część jest zrekonstruowana. Brak zgrubienia kości łuskowej wskazuje na Triceratops a niski róg nosowy jest charakterystyczny dla T. horridus. Ilustracje: LASM, online 2017; 225 Magazine, online 2017. Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (LASM, online 2017);
- (?) USNM (1203, 1206, 1207, 1208, 1209, 1210) - według Hatchera i in. (1907, s. 134) jest to prawie kompletna, dysartykułowana czaszka. Okaz jest zaliczany do Triceratops (Hatcher i in., 1907, s. 134, Lull, 1915, s. 343; Ostrom i Wellnhofer, 1986, s. 118; Lyson i Longrich, 2011), zakrzywiona boczna krawędź kości łuskowej i jej płaska powierzchnia wskazują, że jest to poprawna identyfikacja. Lull (1933, s. 129) oraz Scannella i Horner (2011) podają, że róg nosowy jest słabo wykształcony, zatem jest to prawdopodobnie T. horridus. Ilustracje: Hatcher i in., 1907, tabl. XXXVII; USNM, online. Lokalizacja: formacja Lance (Lyson i Longrich, 2011), hrabstwo Niobrara, Wyoming (Lull, 1933, s. 15);
- WDC LF-001 - niemal kompletny szkielet, zilustrowany i przypisany do T. horridus przez Hartmana [online 2008 - kopia m.in. w Król Padilha, online 2008];

(4) Materiał przypisany na podstawie wieku geologicznego (zob. #Legenda):
- (?) MOR 2552 - brak zgrubienia na kości łuskowej wskazuje, że to Triceratops (Scannella i in., 2014 - macierz), długie rogi nadoczne (ok. 80 cm - Scannella i in., 2014) wykluczają T. prorsus, lecz nie T. ?sp. nov.. Lokalizacja: górne dolne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014; Scannella i in., 2014);
- ^ (?) MOR 2569 - czaszka bardzo młodego osobnika (Goodwin i Horner, 2010), ok. dwukrotnie większa od UCMP 154452. Przednia część czaszki jest znacznie zrekonstruowana (Scannella i in., 2015, s. 233; Farke, online 2014b – zdjęcie z wystawy z MOR). W zmienionej w 2014 r. wystawie w MOR uwzględniona w serii ontogenetycznej T. prorsus (MOR, online 2014). Ilustracje: *; O; Scannella i Horner, 2010, ryc. 3 B; Goodwin i Horner, 2010, ryc. 36.14. Lokalizacja: dolne środkowe warstwy formacji Hell Creek, hrabstwo Garfield, Montana (Goodwin i Horner, 2010; Scannella i Fowler, 2014);
- (?) MOR 2985 - brak zgrubienia na kości łuskowej wskazuje, że to Triceratops (Scannella i in., 2014 - macierz), kość łuskowa mierzy 93 cm (Scannella i in., 2014). Lokalizacja: górne dolne osady formacji Hell Creek, hrabstwo Garfield, Montana (Scannella i Fowler, 2014);
- (?) MOR 8693 - m.in. kość łuskowa o kształcie charakterystycznym dla Triceratops - dość dojrzły okaz (Scannella i in., 2015, ryc. 7.10; s. 448). Lokalizacja: dolne osady formacji Hell Creek, Montana (Scannella i in., 2015);
- (?) okaz z MSP (wcześniej MOR 699) - niemal kompletna czaszka przypisana do Triceratops (Holliday, 2009, zapewne też Farke, 1997) o długości ok. 165 cm, w tym kryza, brak jednak przedniej części pyska i rogów nadocznych (Horner i Goodwin, 2006; Scannella i Fowler, 2014). Ilustracje: "WitmerLab", online; Henderson, online 2012.. Lokalizacja: ?dolne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
(5) Materiał przypisany na podstawie morfologii, wbrew wiekowi geologicznemu (zob. #Legenda):
- UCMP 128561 - holotyp Ugrosaurus olsoni - to fragmenty pyska i kryzy, niezidentyfikowane części czaszki oraz kręg, opisane w CoBabe i Fastovsky (1987). Według Forster (1993) Ugrosaurus to nomen dubium i prawdopodobnie Triceratops (T. sp.) a według Longricha i Fielda (2012) to młodszy synonim Triceratops horridus. Scannella i in. (2014) stwierdzili natomiast, że ze względu na fragmentaryczność jest niejasne, czy okaz reprezentuje morfologię Torosaurus. Fragmenty kryzy mają 2-4 cm grubości (CoBabe i Fastovsky, 1987), co mieści się w zakresie Triceratops, ale nie Torosaurus (zob. opis Torosaurus) - wprawdzie maksymalna grubość przy brzegach kryzy u Torosaurus jest zbliżona a nawet większa niż 4 cm, lecz skoro zachowane fragmenty „wyjawiają niewiele ze struktury i wielkości kryzy” (CoBabe i Fastovsky, 1987, s. 152) a zilustrowana część nie wygląda na obszar jej brzegu, ani wewętrznego ani zewnętrznego (CoBabe i Fastovsky, 1987, ryc. 3.3), to grubość świadczy o przynależności do Triceratops. Budowa rogu nosowego (niskiego, mającego postać guza) może wynikać z różnic międzygatunkowych (Farke, 2011), różnic między osobnikami a nawet z ontogenezy - resorpcji rogu (Scannella i Fowler, 2014) (zob. Zmniejszanie ozdób w ontogenezie dinozaurów: Ceratopsidae). Horner i Goodwin (2008) przedstawili hipotezę, że utrata rogu nosowego za życia zwierzęcia mogła spowodować powstanie chropowatego i nieregularnego guza, jak u Torosaurus MOR 1122 i Ugrosaurus. Róg (epinasale) UCMP 128561 nie został jednak utracony, o czym świadczą brak śladów złamania (CoBabe i Fastovsky, 1987), stwierdzona przez Forster (1993) obecność łukowatego śladu naczynia krwionośnego ("arcuate vessel"), obecnego u Triceratops na przodzie rogu nosowego (zob. Torosaurus: Łukowaty ślad naczynia na rogu nosowym). Również Longrich i Field (2012) oraz Scannella i in. (2014) uznali, że epinasale jest obecny. UCMP 128561 jest prawdopodobnie późnym przedstawicielem T. horridus (zob. dowody kladogenezy). Możliwe jest również, że UCMP 128561 to szczątki dwóch zwierząt - Triceratops (fragmenty kryzy) i Torosaurus (guz nosowy). Scannella i in. (2015, s. 315) podali, że z uwagi na fragmentaryczność „nie wiadomo, czy reprezentuje morfologię Torosaurus”. Status rodzaju Ugrosaurus zależy od ważności Torosaurus latus i Nedoceratops hatcheri. Jeśli Torosaurus i Nedoceratops to młodsze synonimy Triceratops, to Ugrosaurus należy do Triceratops. Jeśli N. hatcheri jest nieważny (jest młodszym synonimem Triceratops horridus), to U. olsoni jest zapewne młodszym synonimem Triceratops. Jeśli jednak Nedoceratops jest ważny, to może on być także jego młodszym synonimem. Wtedy Ugrosaurus olsoni byłby nomen dubium. Jest to jednak mało prawdopodobna hipoteza z uwagi na znacznie większą liczbę odnalezionych T. horridus i tylko jednego Nedoceratops. Szczątki należą do dużego okazu (zob. Torosaurus: Największe ceratopsy). Ilustracje: ** - #Nedoceratops; CoBabe i Fastovsky, 1987; Scannella i Horner, 2011, ryc. 2c. Lokalizacja: górna środkowa część formacji Hell Creek (Scannella i Fowler, 2014; Scannella i in., 2014).
= łącznie 20 osobników przypisanych ponad wszelką możliwość oraz 19 mniej pewnych (=39), w tym 33 niewątpliwie należące do Triceratops.
Lokalizacje
W nawiasach kwadratowych podano liczbę okazów: USA - formacja Lance (Wyoming) [14], formacja Hell Creek (Montana [15], Dakota Południowa [4], Dakota Północna [1]), formacja ?Denver i formacja Laramie (Colorado) [2]; ?Kanada - formacja Frenchman (Saskatchewan) [1].
Etymologia
Horridus to wystający/sterczący (ang. projecting) lub ważny/podniesiony (ang. standing up) (Dodson, 1996).
Galeria
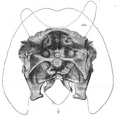
YPM 1820 (holotyp T. horridus) w widoku z tyłu (częściowo zrekonstruowany – por. Tsuihiji, 2010, ryc. 7C). Źródło: Hatcher i in., 1907.
- Hunting dinosaurs in the bad lands of the Red Deer River, Alberta, Canada; a sequel to The life of a fossil hunter (1917) (20765015941).jpg
AMNH 5116. Źródło: Matthew, 1915, ryc. 38
- Triceratops.JPG
BHI 4772 (odlew w Oxford University Museum of Natural History). Fot.: "Ballista".
- Big John triceratops sold at auction in Paris.jpg
"Big John". Fot.: Camille Gévaudan.
- Burpee - Homer the Triceratops 2.JPG
BMRP 2006.4.1. Fot.: Ben Jacobson

DMNH 48617. Fot.: Joey Doll
- Annual report of the Director to the Board of Trustees for the year ... (1901) (18405975546).jpg
FMNH P12003. Źródło: FMNH/Internet Archive Book Images
- Cornudo y Triceratops (11208904415).jpg
HMNS PV.1506. Fot.: Martha Silva
- Triceratops calicornis.jpg
MNHN 1912.20. Fot.: Ignacio García
- Museum of Science, Boston, MA - IMG 3203.JPG
Okaz z MOS. Fot.: "Daderot"

ROM 55380. Fot.: Jeremy Luke Hill

SDSM 2760. Fot.: Paul Weimer
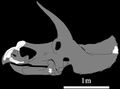
UCMP 128561. Autor: "Laelaps nipponensis"/"GetAwayTrike"

USNM 1205. Źródło: USNM, online

USNM 1201. Źródło: Scannella i Horner, 2011

USNM 2100. Źródło: USNM, online
- Nedoceratops hatcheri 2.jpg
USNM 2412. Fot.: Claire H.

USNM 4928. Źródło: USNM, online









T. sp. nov.
Taksonomia
Osobniki wykopane z górnych warstw środkowej części formacji Hell Creek (ok. 5 metrów poniżej granicy górnych pokładów tej formacji) wykazują cechy pośrednie pomiędzy T. horridus i T. prorsus. Czaszki tych okazów mają m.in. wydłużone rogi nadoczne, charakterystyczne dla triceratopsów z dolnych warstw formacji Hell Creek (T. horridus) oraz wydłużony róg nosowy i głęboki pysk, charakterystyczne dla triceratopsów z górnych warstw (Scannella i in., 2012; 2014) - zob szerzej w Anageneza i heterochronia.
Scannella i in. (2014) nie zaliczyli okazów o pośredniej budowie do żadnego gatunku. Scannella i in. (2015, s. 251-252) uznali je za pośrednie między T. horridus a T. prorsus, wskazując, że dla identyfikacji form z różnych poziomów stratygraficznych potrzebne jest porównanie z holotypami. Możliwe są trzy główne przypisania: 1) do T. horridus, 2) do T. prorsus i 3) do trzeciego gatunku; w grę wchodzić mogą też inne rozwiązania, jak 4) zaliczenie MOR 3045 jako prawdopodobnie bardziej zaawansowanego do T. prorsus a pozostałych okazów do T. horridus, 5) przypisanie wszystkich form do jednego gatunku. Za zaliczeniem okazów pośrednich do T. prorsus przemawia większa ilość cech łączących z tym gatunkiem (sześć - w porównaniu do trzech z T. horridus) i politomia złożona z tych okazów i okazów z górnych warstw (T. prorsus) w wielu z wariantów przeprowadzonej przez Scannellę i in. (2014) analizy kladystycznej. Okazy pośrednie nie wykazują unikalnych cech (autapomorfii), lecz wyłącznie cechy obecne u wcześniejszych i późniejszych form, z wątpliwymi wyjątkami: dużego guza na kości nosowej za rogiem, obecnego u UCMP 113697 i wcześniejszego okazu MOR 2982 (Scannella i in., 2014) oraz ontogenetycznego przekształcenia dość dużego rogu nosowego w guz (obecny u UCMP 128561). Użyteczne wydaje się jednak wydzielenie okazów o pośredniej budowie do osobnego gatunku.
Forster (1996b) uznała T. flabellatus (= Sterrholophus flabellatus, czasem błędnie zapisywany fabellatus) za młodszy synonim T. horridus, za nią Anderson (1999). Longrich i Field (2012, ryc. 5) oraz Longrich (2014) zaliczyli jego holotyp do T. prorsus, zaś Maiorino i in. (2013b) w suplemencie do T. horridus, ale w rycinach widnieje jako T. prorsus. Synonimika z T. prorsus nie została wprost zaproponowana, jednak to T. flabellatus byłby starszym synonimem, gdyż został nazwany rok wcześniej (1889) niż T. prorsus.
Diagnoza
Na podstawie Scannella i in. (2014) cechami pozwalającymi odróżnić T. sp. nov. od pozostałych gatunków (zob. „górne środkowe osady”) (gwiazdką oznaczono cechy obecne u holotypu) są:
(1) krótki pysk - stosunek długości od dzioba do podpórki nosowej kości przedszczękowej (narial strut) powyżej do długości od dzioba do kłykcia potylicznego poniżej 0,28. Obecne także u T. prorsus.
(2) [3] mniej pochylony (bardziej pionowy) wyrostek nosowy kości przedszczękowej - kąt między nim a „rozporą nosową” wynosi powyżej 131 stopni*. Obecne także u T. prorsus.
(3) [14] Szeroki wyrostek nosowy kości przedszczękowej - stosunek wysokości (mierzonej od najniższego punktu artykulacji z kością nosową) do szerokości (długości) najwęższej części wynosi poniżej 1,57*. Obecne także u T. prorsus.
(2-3) obie te cechy, zwłaszcza ta pierwsza, powodowały, że pysk miał opisaną przez Forster (1996b, cecha 4) budowę, tj. „krótki, zaokrąglony pysk”*. Obecne także u T. prorsus.
(5) [22] wyraźny grzbiet na bocznej powierzchni dolnotylnego wyrostka kości przedszczękowej i zawężoną powierzchnię styku z kością nosową. Obecne także u T. prorsus.
(6) [7] dłuższy i wydłużony róg nosowy - ponad 15,5 cm długości u większych okazów (długość od dzioba do kłykcia potylicznego powyżej 1 m). Stopień wydłużenia rogu (długość/szerokość) powyżej 1,6. Długość powyżej 0,15 długości od dzioba do kłykcia potylicznego (u okazów niebędących młodymi, ze słabo rozwiniętymi rogami).
(7-8) [13] guzek na tylnej powierzchni rogu nosowego i [10-11] guzek formowany przez tylną część rogu nosowego i przednią część kości nosowej. Obecne także u T. horridus.
(9) długa kość nosowa – stosunek jej długości do długości od dzioba do kłykcia potylicznego u większych okazów (druga długość ok. 1 m) powyżej 0,38.
(10) [9] prosta dolna krawędź górnej części kości nosowej. Obecne także u T. prorsus.
(11) [12] wyraźny przedniobrzuszny wyrostek kości nosowej. Obecne także u T. horridus.
(12) [1] rogi nadoczne dłuższe niż 0,45 długości od dzioba do kłykcia potylicznego - u większych okazów - o tej długości większej niż 50-90 cm. Obecne także u T. horridus.
Materiał kopalny
(1) Materiał przypisany na podstawie morfologii i wieku geologicznego (zob. #Legenda):
- MOR 3027 ("Yoshi's Trike") - dość kompletny, duży, lecz niedojrzały okaz (Horner i Lamm, 2011) z wielkimi rogami nadocznymi. MOR 3027 jest nieznacznie wcześniejszy (znaleziony ok. 2 m niżej) od MOR 3045 i UCMP 113697. Ilustracje: *; ** - #ontogeneza; MOR, online 2012; Scannella i in., 2014, ryc. S1A. Lokalizacja: górna część środkowych osadów formacji Hell Creek, hrabstwo Garfield, Montana (Scannella i Fowler, 2014);
- MOR 3045 - częściowa czaszka, m.in. kość przedszczękowa, nosowa, róg nadoczny i kryza (Scannella i in., 2014). MOR 3045 jest prawdopodobnie najbardziej zaawansowany z okazów zaliczonych do tego gatunku, o czym świadczyć może (alternatywnie to cechy zmienne w ramach populacji) m.in. przednio-tylnie dłuższy wyrostek nosowy kości przedszczękowej (Scannella i in., 2014).. Ilustracje: Scannella i in., 2014, ryc. S1B. Lokalizacja: górna część środkowych osadów formacji Hell Creek, hrabstwo Garfield, Montana (Scannella i Fowler, 2014);
- UCMP 113697 - zanim rozpoznano, że to okaz T. ?sp. nov., określenie przynależności gatunkowej było sporne - róg nosowy wykazuje budowę typową dla T. prorsus (Forster, 1996b), lecz ma bardzo długie rogi nadoczne (82 cm - Horner i Goodwin, 2006), co jest cechą T. horridus (Forster, 1996b). Forster (1996b), Anderson (1999) oraz Lyson i Longrich (2011) zaliczyli go do T. horridus a Longrich i Field (2012) do T. prorsus. Ilustracje: *; ** - #Długość rogów nadocznych; O; Ford, online – Triceratops; Scannella i in., 2014, ryc. 1I]. Lokalizacja: górne środkowe warstwy formacji Hell Creek, hrabstwo McCone, Montana (Forster, 1996b; Scannella i Fowler, 2014).
Z tego samego przedziału stratygraficznego pochodzi także UCMP 128561 (Scannella i Fowler, 2014) - holotyp Ugrosaurus, lecz wykazuje budowę rogu (guza) nosowego wcześniejszych T. horridus.
(2) Materiał nieprzypisany w literaturze (zob. #Legenda):
- SMM P62/1/1 (zapisywany również SMM P62.1.1) - jest zmontowany wraz z nieco większym SMM P60/2/1 w szkielet wystawiony w SMM (Erickson, 1966, s. 3 podaje, że na potrzeby uzupełnienia okazu P62/1/1 m.in. lewa kość jarzmowa została zastąpiona przedziurawionym elementem SMM P60/2/1 i na s. 4, że w P62/1/1 kość przedzębowa była zgnieciona i została wstawiona pochodząca od innego okazu). Wszystkie badania posługują się tylko jednym z tych okazów (P62/1/1: Forster, 1990; 1996a; 1996b; Farke, 1997; 2004; Anderson, 1999; P60/2/1: Scannella i Horner, 2010; Scannella i in., 2015; numer P60.1 jest widoczny w tym ostatnim badaniu na ryc. 7A). Jest to wielki okaz - długość całkowita szkieletu to 7,92 m, kręgosłupa 6,68 m, czaszki 2,21 m, od dzioba do kłykcia potylicznego 1,27-1,31 m (Anderson, 1999; Erickson, 1966) - wymiary te są przeszacowane, gdyż dziób i ogon zostały zrekonstruowane jako zbyt długie (zob. też Torosaurus: Największe ceratopsy). Przednia część pyska nie zachowała się, jak również kości przedszczękowe, części okolicy nosowej za nozdrzem i wiele z okolicy kości szczękowej, a także niektóre części przedniej krawędzi kości ciemieniowej (Erickson, 1966). SMM P62/1/1 został przez Ericksona zaliczony do T. prorsus a później do T. horridus przez Forster (1996b). Forster wskazała, że SMM P62/1/1 łączy cechy T. horridus: obecne też u T. ?sp. nov. długie rogi nadoczne (72 cm - Forster, 1990, s. 164) i otwarte ciemiączko ciemieniowo-czołowe (Erickson, 1966, s. 4, podał, że ciemiączko jest częściowo zamknięte – otwór jest zredukowany [ang. bridged over] jedynie w centrum), oraz T. prorsus: obecny też u T. ?sp. nov. duży róg nosowy o długości 27 cm (Erickson, 1966). Dolna krawędź górnej części kości nosowej jest dość prosta, co sugeruje, że jest to T. prorsus lub T. ?sp. nov.. Różki na brzegu kryzy są (choć słabo) przyrośnięte (Erickson, 1966) a powierzchnia tylnej części kryzy jest chropowata, co wskazuje, że okaz osiągnął co najmniej zaawansowane stadium rozwoju według Longricha i Fielda (2012) i jest dojrzalszy niż niemające ww. cech i mające otwarte ciemiączka T. ?sp. nov. UCMP 113697 i T. prorsus GMNH-PV 124. Należałoby się spodziewać, że w tej fazie T. prorsus powinien mieć już zamknięte ciemiączko - choć nie jest znane, w którym z etapów ontogenezy wyróżnionych przez Longricha i Fielda (2012) ciemiączko zarastało, wydaje się mało prawdopodobne, aby dopiero w ostatniej, skoro Torosaurus ANSP 15192, który jest w przedostatniej fazie (Longrich i Field, 2012), ma je zamknięte (Farke, 2006). Taka kombinacja cech sugeruje T. ?sp. nov.. Ilustracje: *; Erickson, 1966. Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (Forster, 1996b; Erickson, 1966).
= łącznie 4 osobniki przypisane ponad wszelką możliwość.
Galeria
- Yoshi-Trike-0006-black01.jpg
MOR 3027. Fot.: "WernerG2011"
- Triceratops Science Museum MN.JPG
SMM P62/1/1. Fot.: Nielseno
- UCMP Triceratops left.JPG
UCMP 113697. Fot.: "EncycloPetey"
T. prorsus
Diagnoza
Na podstawie Scannella i in. (2014) cechami pozwalającymi odróżnić T. prorsus od pozostałych gatunków (zob. „górne osady”) (gwiazdką oznaczono cechy obecne u holotypu) są:
(1) krótki pysk - stosunek długości od dzioba do podpórki nosowej kości przedszczękowej (narial strut) powyżej do długości od dzioba do kłykcia potylicznego poniżej 0,28. Obecne także u T. ?sp. nov..
(2) [3] mniej pochylony (bardziej pionowy) wyrostek nosowy kości przedszczękowej - kąt między nim a „rozporą nosową” wynosi powyżej 131 stopni. Obecne także u T. ?sp. nov., do wartości 142 stopni. Cecha ta powodowała, że pysk miał opisaną przez Forster (1996b, cecha 4) budowę, tj. „krótki, zaokrąglony pysk”*.
(6) [7] dłuższy i wydłużony róg nosowy - ponad 15,5 cm długości u większych okazów (długość od dzioba do kłykcia potylicznego powyżej 1 m). Stopień wydłużenia rogu (długość/szerokość) powyżej 2 i często powyżej 2,2*. Długość rogu powyżej 0,2 długości od dzioba do kłykcia potylicznego (u okazów niebędących młodymi, ze słabo rozwiniętymi rogami)*.
(7-8) [13, 10-11] zredukowany (lub brak) guz na górnej powierzchni rogu nosowego i zredukowany lub nieobecny guzek formowany przez tylną część rogu nosowego i przednią część kości nosowej.
(9) długa kość nosowa – stosunek jej długości do długości od dzioba do kłykcia potylicznego u większych okazów (druga długość ok. 1,1 m) powyżej 0,41.
(12) [1] rogi nadoczne długości do 0,64 długości od dzioba do kłykcia potylicznego i często krótsze niż 0,45 długości podstawowej czaszki (u większych okazów o tej długości większej niż 50-90 cm) - obecne też u Torosaurus latus.
Materiał kopalny
Holotyp to YPM 1822 - niemal kompletna czaszka (zrekonstruowana została tylna część kryzy) i kilka (6-7) kręgów szyjnych, w tym syncervical (Marsh, 1890a; Hatcher i in., 1907). Jest to najmniejszy dorosły okaz Triceratops (Longrich i Field, 2012) (nie licząc problematycznego Tatankaceratops). Czaszka ma 152,3/153,2/154,9 cm (Hatcher i in., 1907; Lull, 1933; Anderson, 1999) a od dzioba do kłykcia potylicznego 88 cm (Forster, 1996b; Anderson, 1999) lub 93 cm (Borkovic, 2013). Okaz opisany w: Hatcher i in., 1907, s. 127 i n. Ilustracje: ** - #Kryza = Hatcher i in., 1907, tabl. XXXII-XXXVI, ** = Longrich i Field, 2012, ryc. 1a; Forster, 1996a, ryc. 7. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Hatcher i in., 1907; Ostrom i Wellnhofer, 1986).


(1) Materiał przypisany na podstawie morfologii i wieku geologicznego (zob. #Legenda):
- ESU 2014-1 - niekompletna czaszka młodego osobnika. Kość ciemieniowa z 5 episquamosales (Roberts, 2019) sugeruje Triceratops a prosta krawędź kości nosowej T. prorsus. Ilustracje: Roberts, 2019, ryc. 17. Lokalizacja: górne warstwy formacji Hell Creek, hrabstwo Garfield, Montana (Roberts, 2019);
- MWC 7584 (= BYU 12183 - MWC, online 2011a; w suplemencie publikacji Scannelli i in. [2014] oznaczony MWC 8574) – jedna z największych czaszek Triceratops (zob. Torosaurus: Największe ceratopsy), mierząca 2,5 m (Scannella i Horner, 2010), ok. 2,7 m (Olshevsky w "Dino Frontline, 6", za "Laelaps nipponensis"/"GetAwayTrike", prywatna korespondencja, I.2014), ok. 2,74 m (=9 stóp; MWC, online 2011b) lub 2,89 m i 1,95 m szerokości (Curtice, online 2021). Długość podstawowa czaszki (basal skull length) to ok. 1,2 m (Farke, online 2004) ok. 1,386 m (Scannella i in., 2014, oszacowanie na podstawie długości kości jarzmowej) lub ok. 1,5 m (Olshevsky w "Dino Frontline, 6", za "Laelaps nipponensis"/"GetAwayTrike", prywatna korespondencja, I.2014). Ma zrekonstruowane kluczowe obszary w kryzie, więc nie jest pewne, czy ma okna (Beach, 2011a; 2011b). Wydaje się jednak (zob. zdjęcia na Dinosaur Journey Museum, online 2011), że to w istocie Triceratops - po lewej stronie czaszka ma zachowane ok. 2/3 kryzy, która nie wykazuje budowy Torosaurus - prawdopodobnie nie ma podłużnego wału na kości łuskowej wzdłuż łączenia z kością ciemieniową (ang. squamosal bar). Róg nosowy jest - nie uwzględniając problematycznego duranteceratops - najdłuższy z odnotowanych u Triceratops (36,6 cm - Scannella i in., 2014). Ilustracje: *; MWC, online 2011a; Clemens i Hartman, 2014, ryc. 43. Lokalizacja: górne górne osady formacji Hell Creek, Montana (MWC, online 2011b; Farke, online 2004; Scannella i in., 2014);
- MOR 004 ("MORT") – prawie kompletna (górne części rogów nadocznych są zrekonstruowane - Scannella i Fowler, 2014, s. 327, contra Horner i Goodwin, 2006) dość duża czaszka (dł. całkowita 208 cm [Horner i Goodwin, 2006], podstawowa 122 cm [Scannella i in. 2014]; Horner i Goodwin [2006] podają 94 cm, lecz długości podawane przez tych autorów są znacznie niższe niż podawane przez Scannellę i in. [2014] i prowadzą do wniosku, że mierzone przez nich okazy wyróżniają się na tle wszystkich innych nietypowymi proporcjami: długość podstawowa czaszki byłaby znacznie niższa w porównaniu do całkowitej niż u innych okazów, co sugeruje, że podane pomiary to nie długość od początku dzioba do końca kłykcia potylicznego). Długi (28,5 cm - Scannella i in., 2014), wydłużony i zwrócony do przodu róg nosowy wskazuje, że to T. prorsus. Ilustracje: *; ** - Długość rogów nadocznych; Materiał kopalny ogólnie; Horner i Goodwin, 2006, ryc. 1e; Barton, online 2004; Scannella i in., 2014, ryc. 1H. Lokalizacja: górne górne osady formacji Hell Creek, Montana (Horner i in., 2011; Scannella i Fowler, 2014);
- ^ $ MOR 598 - izolowany róg nosowy, którego kształt sugeruje T. prorsus (Scannella i in., 2015, ryc. 7.7A). Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 1110 - czaszka młodego osobnika o długośći całkowitej 135 cm i podstawowej 77,6 cm. Według Hornera i Goodwina (2006) jest kompletna, lecz Scannella i in. (2014, suplement) podają, że dziób został zrekonstruowany (to samo wynika z informacji na wystawie MOR – Tanke, online 2014). MOR 1110 został zaliczony do T. prorsus przez Longricha i Fielda (2012). Maiorino i in. (2013a; 2013b) w suplementach zaliczyli go do T. horridus, lecz na rycinach oznaczono go jako T. prorsus. Ilustracje: ** - #Anageneza i heterochronia; O; Horner i Goodwin, 2006, ryc. 1c. Lokalizacja: górne osady formacji Hell Creek, Montana (Horner i in., 2011);
- MOR 1625 - mniej niż 6 epiparietales i episquamosales i brak zgrubienia na kości łuskowej wskazuje, że to Triceratops (Scannella i in., 2014 - macierz), zaś m.in. wydłużony i duży róg nosowy (30 cm – Scannella i in., 2014) wskazuje, że to T. prorsus. Rozmiary kłykcia potylicznego wskazują, że to wielki okaz (zob. Torosaurus: Największe ceratopsy). Ilustracje: Horner i Goodwin, 2008, ryc. 4c]. Lokalizacja: dolne górne osady formacji Hell Creek, Montana (Horner i in., 2011; Scannella i Fowler, 2014);
- MOR 2574 - brak zgrubienia na kości łuskowej wskazuje na Triceratops a wydłużony róg nosowy (Scannella i in., 2014) na T. prorsus. Okaz jest niedorosły, o czym świadczą zwrócone ku tyłowi końce rogów nadocznych (Keenan i Scannella, 2014). Ilustracje: Horner i Goodwin, 2008, ryc. 1F, 2C; Horner w YPM, online 2013; Keenan i Scannella, 2014; Scannella i in., 2014, ryc. S2F. Lokalizacja: dolne górne osady formacji Hell Creek, hrabstwo Garfield, Montana (Horner i in., 2011; Scannella i Fowler, 2014);
- ^ $ MOR 2590 - częściowa kość przedszczękowa (Scannella i Fowler, 2014), szeroki kąt między nosowym wyrostkiem kości przedszczękowej a "podpórką" nosową (narial strut) sugeruje, że to T. prorsus (kąt jest nieznacznie szerszy niż u T. ?sp. nov.) (Scannella i Fowler, 2014). Lokalizacja: ?górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014; Scannella i in., 2014);
- MOR 2702 - duży okaz - m.in. kość łuskowa bez zgrubienia o długości 100 cm, niekompletna kość ciemieniowa (grubość od 3 do 0,6 cm), kość nosowa wraz z wydłużonym i długim rogiem mierzącym 30 cm, rogi nadoczne (dł. ok. 55 cm) i kość udowa; żuchwa o dł. ok. 60 cm sugeruje, że długość podstawowa czaszki to ok. 121 cm (Keenan i Scannella, 2014; Scannella i in., 2014). Ilustracje: Scannella i Horner, 2010, ryc. 3G; Keenan i Scannella, 2014; Scannella i in., 2014, ryc. S2G. Lokalizacja: dolne górne osady formacji Hell Creek, hrabstwo Garfield, Montana (Horner i in., 2011; Keenan i Scannella, 2014);
- MOR 2923 ("Joe's Trike") - okaz zidentyfikowany jako T. prorsus (MOR, online 2013) i ma wydłużony i duży (29 cm - Scannella i in., 2014), skierowany do przodu róg nosowy i zamknięte ciemiączka czołowo-ciemieniowe. Ilustracje: *; Scannella i Fowler, 2014, s. 319; MOR, online 2013. Lokalizacja: górne górne warstwy formacji Hell Creek, hrabstwo Garfield, Montana (MOR, online 2013; Scannella i Fowler, 2014);
- ^ MOR 2924 - niemal kompletna czaszka młodego okazu, w tym kość ciemieniowa, elementy pozaczaszkowe (Scannella i Fowler, 2014), wypukła kość łuskowa i brak zgrubienia na niej oraz brak okien w kryzie wskazują, że to Triceratops (Scannella i Horner, 2010, ryc. 1B; Scannella i in., 2014 – macierz), zaś wydłużony róg nosowy i pionowy wyrostek nosowy kości przedszczękowej, że to T. prorsus (Scannella i in., 2014; Scannella i in., 2015, ryc. 7.7A). Ilustracje: Scannella i Horner, 2010, ryc. 1B; Scannella i in., 2014, ryc. S4C. Lokalizacja: dolne górne osady formacji Hell Creek, hrabstwo Garfield, Montana (Scannella i Fowler, 2014);
- $ MOR 2936 - zawiera m.in. kość łuskową (Scannella i Fowler, 2014), brak guzka na tylnej powierzchni rogu nosowego i przednio-tylnie długi wyrostek nosowy kości przedszczękowej wskazują, że to T. prorsus (Scannella i in., 2014). Lokalizacja: dolne górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- $ MOR 2938 - m.in. niekompletna kość ciemieniowa (Scannella i Fowler, 2014), wydłużony i długi na 29 cm róg nosowy wskazuje, że to T. prorsus (Scannella i in., 2014). Lokalizacja: górne górne osady formacji Hell Creek, hrabstwo Garfield, Montana (Scannella i Fowler, 2014; Scannella i in., 2014);
- $ MOR 2941 - pazur i wydłużony, długi na 24,8 cm róg nosowy, który wskazuje, że to T. prorsus (Scannella i in., 2014). Lokalizacja: górne górne osady formacji Hell Creek, Montana (Scannella i in., 2014);
- MOR 2951 ("DF Juvie Trike 3") - najbardziej kompletny z młodych osobników (Scannella i Fowler, 2014) - niemal kompletna czaszka o długości podstawowej 58,6 cm (Scannella i in., 2014), część szkieletu pozaczaszkowego (Scannella i Fowler, 2014). Dość przednio-tylnie długi wyrostek nosowy kości przedszczękowej (dłuższy niż u T. horridus, lecz krótki, jak na T. ?sp. nov. i T. prorsus) i szeroka tylnobrzuszna powierzchnia dolnotylnego wyrostka kości przedszczękowej wskazują, że to T. prorsus (Scannella i in., 2014). Ilustracje: ** - #Media, T. ?sp. nov., Ontogeneza; O; Scannella i Horner, 2010, ryc. 1A. Lokalizacja: dolne górne osady formacji Hell Creek, Montana (Horner i in., 2011; Scannella i Fowler, 2014);
- $ MOR 2952 - okaz zawiera m.in. fragmenty kryzy (Scannella i Fowler, 2014), długi na ponad 27 cm i wydłużony róg nosowy wskazuje, że to T. prorsus; kształt jest zbieżny z T. ?sp. nov. i T. prorsus (Scannella i in., 2015, ryc. 7.7A). Rogi naodoczne mają ok. 60 cm (Scannella i in., 2014). Lokalizacja: górne górne warstwy formacji Hell Creek, Montana (Horner i in., 2011; Scannella i Fowler, 2014; Scannella i in., 2014);
- ^ $ MOR 2965 - izolowany róg nosowy, którego wydłużenie i kształt sugeruje T. prorsus (Scannella i in., 2014; Scannella i in., 2015, ryc. 7.7A). Lokalizacja: górne górne osady formacji Hell Creek, Montana (Scannella i in., 2014);
- MOR 2971 - duża czaszka zawierająca m.in. kryzę, kość przedszczękową, dziób i róg nosowy (Scannella i Fowler, 2014). Położone daleko od siebie otwory na brzusznej powierzchni kości przedszczękowej (Scannella i in., 2014 – macierz, cecha 20) sugerują, że to Triceratops (zob. Torosaurus: Budowa kości przedszczękowej), zaś skierowany pionowo i przednio-tylnie długi wyrostek nosowy kości przedszczękowej, długi róg nosowy, szeroki kąt między nosowym wyrostkiem kości przedszczękowej a "podpórką" nosową (narial strut) i brak guzka na górnej powierzchni kości nosowej, wskazują, że to T. prorsus (Scannella i in., 2014). Lokalizacja: dolne górne osady formacji Hell Creek, Montana (Horner i in., 2011; Scannella i Fowler, 2014);
- $ MOR 2972 - część kości łuskowej, mózgoczaszka i inne elementy (Scannella i Fowler, 2014). Niezwykłą cechą, przywodzącą na myśl Tatankaceratops, jest zbliżona długość wszyskich rogów - róg nadoczny jest wydłużony i mierzy 26,7 cm (w porównaniu do długości podstawowej czaszki ok. 98 cm, daje to stosunek 0,27 - Scannella i in., 2014) a róg nosowy 25 cm (Scannella i in., 2014), co wskazuje, że to T. prorsus. Lokalizacja: dolne górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 2978 - czaszka nie zawierająca kości przedszczękowej, szczękowej i żuchwy (Scannella i Fowler, 2014), wypukła kość łuskowa i brak zgrubienia na niej oraz brak okien w kryzie wskazują, że to Triceratops (Scannella i in., 2014 - macierz), zaś m.in. wydłużony róg nosowy (Scannella i in., 2014), wydłużona i prosta kość nosowa, krótkie rogi nadoczne i zamknięte ciemiączko czołowo-ciemieniowe (Scannella i in., 2014 - macierz), że T. prorsus (Scannella i in., 2014). Ilustracje: *; MOR, online 2019a. Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i in., 2014);
- MOR 2999 - m.in. kość ciemieniowa, łuskowa, nosowa, rogi nadoczne i mózgoczaszka. Brak zgrubienia kości łuskowej wskazuje na Triceratops a zredukowany przedniobrzuszny wyrostek kości nosowej na T. prorsus (Scannella i in., 2014, cecha 12). Ilustracje: MOR, online 2014; Harper, online 2014; Scannella i in., 2015, ryc. 7.15.C. Lokalizacja: górne górne warstwy formacji Hell Creek, hrabstwo Garfield, Montana (Scannella i Fowler, 2014);
- $ MOR 3008 - izolowany długi (26,5 cm) i wydłużony róg nosowy wskazuje przynależność do T. prorsus (Scannella i Fowler, 2014; Scannella i in., 2014). Lokalizacja: dolne górne osady formacji Hell Creek, hrabstwo Garfield, Montana (Scannella i Fowler, 2014; Scannella i in., 2014);
- ^ $ MOR 3048 - m.in. wydłużony róg nosowy sugerujący T. prorsus (Scannella i in., 2014) o kształcie zbieżnym z T. ?sp. nov. i T. prorsus (Scannella i in., 2015, ryc. 7.7A) i kłykieć potyliczny (Scannella i Fowler, 2014). Lokalizacja: górne górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- ^ $ MOR 3073 - róg nosowy, którego kształt sugeruje T. prorsus (Scannella i in., 2015, ryc. 7.7A). Róg jest wystawiony obok MOR 1110 i na podstawie porównania ich rozmiarów wydaje się, że MOR 3073 ma ponad 20 cm długości. Ilustracje: Evanston, online 2015. Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i in., 2015, s. 450);
- $ SUP 9713 (również jako 9713.0) - zawiera kość łuskową i fragmenty ciemieniowej (Happ, 2008). Ma róg nosowy o długości 30 cm (wraz z kością nosową), co sugeruje T. ?sp. nov. lub T. prorsus. Prawy róg nadoczny ma 60 (Happ, 2008) lub 71,8 cm (Happ, 2010). Okaz ma ślady po walce z Tyrannosaurus (Happ, 2008), oraz tkankę miękką z rogu nadocznego i mózgoczaszki (Happ i Morrow, 2000; Happ, 2010) (pozostałości po tkance rogowej znajdowały się także na rogu nadocznym YPM 1821 - Hatcher i in., 1907, s. 32 oraz według niepublikowanych danych w rogu nadocznym okazu z Hell Creek - Morales i Campbell, 2006). Ilustracje: Happ, 2008; 2010. Lokalizacja: górne warstwy formacji Hell Creek, okolice Jordan, hrabstwo Garfield, Montana (Happ, 2010);
- UCMP 136092 - dość kompletna czaszka o długości ok. 160 cm i budowie Triceratops (Goodwin i Horner, 2010). Pysk nie zachował się, lecz bardzo krótkie rogi nadoczne (zbliżające go do [[Tatankaceratops) sugerują, że to T. prorsus. Ilustracje: Goodwin i Horner, 2010, ryc. 36.11; Scannella i Fowler, 2014, ryc. 3; KCJ44, online 2014. Lokalizacja: górne górne osady formacji Hell Creek, hrabstwo McCone, Montana (Goodwin i Horner, 2010; Scannella i Fowler, 2014);
- UND 3000 - czaszka przypisana do T. prorsus przez Hollanda (1997). Choć kryza przynajmniej w pewnych miejscach jest cienka (poniżej 1 cm) i w dużej części niekompletna, zachowane fragmenty wskazują, że miał prawdopodobnie 15 różków na jej obrzeżu (Holland, 1997), co wskazuje, że to Triceratops, zaś długi (ok. 25-30 cm; z pewnością nie 45 cm, które podał Holland) i wydłużony róg nosowy (Holland, 1997) dowodzi, że to T. prorsus. Bardzo długie (84 i 78 cm) rogi nadoczne zostały zrekonstruowane (Holland, 1997). Czaszka opisana w: Holland, 1997. Ilustracje: *; Holland, 1997. Lokalizacja: górne warstwy formacji Hell Creek, hrabstwo Slope, Dakota Północna (Holland, 1997; Pearson i in., 2002).


(2) Materiał przypisany w literaturze na podstawie morfologii (niektóre mogą reprezentować T. ?sp. nov.) (zob. #Legenda): - Tokaryk, 1986:
- $ EM P15.1 – gigantyczna czaszka (zob. Torosaurus: Największe ceratopsy), zaliczona do T. prorsus (Tokaryk, 1986), ale niezawierająca kryzy, więc nie jest pewne, czy to Triceratops. Róg nosowy jest bardzo długi (34,6 cm wzdłuż przedniej krawędzi - Tokaryk, 1986), jak u T. prorsus. Czaszka jest bardziej fragmentaryczna niż wynika z opisu Tokaryka, według którego okolica kości jarzmowej, kwadratowo-jarzmowej, kwadratowej została nieprawidłowo zrekonstruowana. Jak podała Bamforth (online 2019b), „fragmenty czaszki zostały sklejone, brakujące elementy uzupełniono i całość pomalowano. Obecnie ponownie preparujemy okaz, by sprawdzić, jaka część jest rzeczywiście zachowana”. Z fotografii (Bamforth, online 2019b; Cypress Hills-Grasslands Destination Area, online 2019) wynika, że na ewentualny zbyt duży rozmiar okazu wpływać może jedynie kilkucentymetrowy fragment uzupełniony między kością dziobową a nosową. Ilustracje: *; Tokaryk, 1986, ryc. 2; EM, online 2012; The Boomtown, online 2013; Bamforth, online 2019b; Cypress Hills-Grasslands Destination Area, online 2019. Lokalizacja: formacja Frenchman, dolina rzeki Frenchman na południe od Shaunavon, Saskatchewan (Tokaryk, 1986);
- SMNH P1163.4 – czaszka i prawdopodobnie żebra i elementy miednicy (Tokaryk, 2009), przypisana przez Tokaryka (1986) do T. prorsus. Długi na ponad 31,5 cm róg nosowy potwierdza klasyfikację. Ilustracje: *; Tokaryk, 1986, ryc. 1. Lokalizacja: formacja Frenchman, dolina rzeki Frenchman na południe od Shaunavon, Saskatchewan (Tokaryk, 1986);
- Forster, 1996b:
- BSP 1964 I 456 (wcześniej YPM 1834) - holotyp T. brevicornus (Hatcher, 1905), czasem błędnie zapisywany brevicornis lub przypisywany Lullowi. Okaz jest uniwersalnie uznawany za T. prorsus - prócz Maiorino i in. (2013a). Okaz opisany w: Hatcher i in., 1907, s. 141-142 (czaszka) i 46-51 (kręgi) oraz w Ostrom i Wellnhofer, 1986. Ilustracje: *; Hatcher i in., 1907, tabl. XLI-XLII; Ostrom i Wellnhofer, 1986, zwłaszcza ryc. 3 i tabl. I. Lokalizacja: późniejsze osady formacji Lance („blisko szczytu Laramie”), hrabstwo Niobrara, Wyoming (Hatcher i in., 1907; Ostrom i Wellnhofer, 1986);
- CM 1219 (pomyłkowo podpisany CM 1221 w Forster, 1993 i 1996b; różnice w wyglądzie między 1219 a ilustracjami Forster wynikają z dodatkowej preparacji w 2007-2008 r. - M. Lamanna, prywatna korespondencja, I.2014) - okaz opisany przez Lulla (1933, s. 119-120) jako T. brevicornus, ma od jego holotypu większy i masywniejszy róg nosowy; brak ciemiączka ciemieniowo-czołowego wskazuje, że to T. prorsus (Forster, 1996b). Ilustracje: *; Glut, 1997, s. 17; McIntosh, 1981, ryc. 22. Lokalizacja: formacja Hell Creek, hrabstro Garfield, Montana (Lull, 1933, s. 119-120);
- LACM 59049 – czaszka mierząca „ponad 8 stóp (244 cm)” i 60% szkieletu (Stock, online 2015b), w tym żebra i kręgi (LACM, online). Krótki pysk i prosta dolna krawędź górnej części kości nosowej wskazują, że to T. prorsus. W literaturze często określa się go jako 7207 (Forster, 1996b; Lehman, 1998; Wu i in., 2007; Farke, 2011; na ryc. 7 w Forster [1996b] podpisany jako LACM 7202, lecz jest to nazwa lokalizacji LACM 27428 [LACM, online]), lecz jest to numer lokalizacji (Samuel McLeod, LACM, prywatna korespondencja, I.2014). Numer 59049 podają Scannella i Horner (2010) oraz Maiorino i in. (2013b). W tabeli Andersona (1999) są one osobno i różnią się wysokością kłykcia potylicznego. Glut (2006, s. 221) podpisał czaszkę jako 7207/59049. W LACM znajduje się także okaz będący odlewem LACM 59049 (Glut, 2000, s. 355; Stock, online 2015a), widoczne są jednak pewne różnice (m.in. w przedniej dolnej części kości przedszczękowej, nietypowy jest kształt epiparietales i episquamosales) w porównaniu do ryc. 3B w Forster, 1996b i z pewnością przedstawiającego ten sam okaz zdjęcia (DeLuca, online 2011). Ilustracje: ** - #Media; Forster, 1996b, ryc. 3b; Glut, 1997, s. 65; 2006, s. 221; DeLuca, online 2011; Shore, online 2015. Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (Forster, 1996b; LACM, online);
- Sampson i in., 2010:
- ^ ? LACM 7303;
- Longrich i Field, 2012:
- GMNH-PV 124 - dość kompletna czaszka, kręgi przedkrzyżowe (brak jednego), kość krzyżowa, 12 kręgów ogonowych, żebra i górne części kończyn. Długi na ponad 20 cm i wąski róg nosowy (stosunek długości do wysokości to ok. 2,65 – jedna z najwyższych wartości u Triceratops – za ryc. 4 w Fujiwara i Takakuwa, 2011) wskazują, że to T. prorsus. Kąt między wyrostkiem nosowym kości przedszczękowej a „rozporą nosową” wynosi 128-134 stopni (ryc. 2A w Fujiwara i Takakuwa, 2011), zatem jest pośredni między T. horridus a zaawansowanymi Triceratops (por. Scannella i in., 2014). Wyrostek nosowy wydaje się jednak bardzo szeroki (stosunek wysokości do szerokości ok. 1,08), co znamionuje T. prorsus. Wizualnie budowa wyrostka również odpowiada T. prorsus lub zaawansowanemu MOR 3045 (Scannella i in., 2014, ryc. S2). GMNH-PV 124 został zaliczony do T. prorsus przez Longricha i Fielda (2012) oraz Longricha (2014). Okaz opisany w: Fujiwara i Takakuwa, 2011. Ilustracje: Fujiwara i Takakuwa, 2011. Lokalizacja: formacja Hell Creek, hrabstwo Harding, Dakota Południowa (Fujiwara i Takakuwa, 2011);
- (?) OMNH 10170 – gruba na 3,5 cm kryza (Longrich, 2011) sugeruje, że to prawdopodobnie Triceratops (Scannella i Horner, 2010), okaz zawiera kości ciemieniową, łuskową, nosową, przedszczękową i róg nosowy (Longrich, 2011, Longrich i Field, 2012 – kodowania). Został zaliczony do T. prorsus przez Longricha i Fielda (2012) oraz Longricha (2014);
- Maiorino i in., 2013a/2013b:
- TCM 2004.49.1 - czaszka dorosłego osobnika (Maiorino i in., 2013b). Jest to zapewne okaz ilustrowany obok, gdyż budowa i miejsce pochodzenia jest zbieżne z danymi Maiorino i in. a brak danych o trzecim okazie z TCM. Brak zgrubienia kości łuskowej wskazuje na Triceratops a wydłużony róg nosowy na T. prorsus. Ilustracje: *; Evans, online 2014; Hall, online 2016. Lokalizacja: formacja Hell Creek, Dakota Południowa (Maiorino i in., 2013b/Howie, online 2010);
- ^ (?) BHI 6409 - zachowana jest przynajmniej kość łuskowa, według badań morfometrycznych należąca do Triceratops (Maiorino i in., 2013b). Lokalizacja: formacja Hell Creek, Dakota Południowa (Maiorino i in., 2013b);
- LACM 151459 - czaszka, szyja i kończyna przednia, zmontowana w szkielet z kilkoma innymi okazami (LACM - za Mallison, online 2013). Krótki pysk wskazuje na T. prorsus. Ilustracje: *; ** - Materiał kopalny ogólnie. Lokalizacja: formacja Hell Creek, hrabstwo Carter, Montana (Maiorino i in., 2013b; LACM, online).

.jpg)
(3) Materiał nieprzypisany w literaturze (niektóre mogą reprezentować T. ?sp. nov.) (zob. #Legenda):
- BSM 1958G00003.00001 (numer za Mabbett, online 2015; =GLAHM 152339 [odlew]; =AMNH 5028) - kompletna czaszka (AMNH, online – 5028) mierząca 210 cm (Lull, 1933, s. 117). Okaz został wzmiankowany w literaturze chyba jedynie przez Lulla (1933, s. 117), który zaliczył go do T. horridus. Jednak jego opisu wynika, że to T. prorsus: okaz ma róg nosowy większy niż u T. brevicornus i skierowany do przodu (kończący się tuż przed koniuszkiem dzioba) oraz krótsze niż u YPM 1820 i YPM 1822 rogi nadoczne. Nietypowymi cechami są brak wyraźnej strefy ze śladami naczyń krwionośnych na dolnej krawędzi kryzy oraz nieproporcjonalnie duża przednia część czaszki (większa niż u YPM 1820) a mała kryza (Lull, 1933), która sugeruje, że to Triceratops. Według informacji GLAHM (online 2017), znajdujący się w nim odlew to Triceratops odkryty w 1908 roku w Montanie (formacja Hell Creek) przez ekipę składającą się z m.in. Browna, Kaisena i Lamberta z AMNH (ponadto obecnie miejsce odkrycia jest zalane przez Fort Peck Reservoir). Czaszka została zakupiona przez „BM&AG” (zapewne Birmingham Museum & Art Gallery) w 1958 r. a w 2000 r. została przekazana do Thinktank (BSM) i tam wystawiona. Również według BSM (online) okaz odkopano w 1908 roku w Montanie. Informacje te pokrywają się z danymi podanymi przez Lulla (1933, s. 117) o odkryciu czaszki przez Browna w 1908 r. w hrabstwie Dawson w Montanie (formacja Hell Creek). W GLAHM znajduje (znajdował?) się odlew o szarym kolorze, bardzo podobny do okazu z BSM o brązowym kolorze i okazu widocznego na zdjęciach AMNH (por. odpowiednio: Shetland Museum and Archives, online oraz GLAHM, online 2017; "ALEXCOTTERELL", online 2014; AMNH, online). Widoczna na zdjęciach budowa odpowiada opisowi Lulla. Ilustracje: w BSM: ** - #Media; "ALEXCOTTERELL", online 2014; BSM, online; w GLAHM: Shetland Museum and Archives, online; GLAHM, online 2017; w AMNH: AMNH, online. Lokalizacja: formacja Hell Creek, hrabstwo Dawson/Garfield, Montana (Lull, 1933; AMNH, online – 5028);
- ^ $ okaz z CCM - Wolff (2007) wylicza dwa okazy z CCM: "Full Mount" oraz "Half mount" (jednym z nich jest zapewne CCM 49-1). Na zdjęciach Johnsona (online 2014b) widoczny jest okaz z CCM z zachowaną kryzą oraz długim rogiem nadocznym (wydaje się większy niż u T. ?sp. nov.) a krótkimi nadocznymi. Ilustracje: Johnson, online 2014b;
- (?) okaz z DMC ("Bill") - praktycznie kompletna czaszka o długości ok. 193 cm (=76 cali) z raczej krótkim pyskiem i dużym rogiem nosowym (DMC, online a). Ilustracje: *; Tanke, online 2016. Lokalizacja: formacja Hell Creek, Montana (DMC, online a);
- ? KUVP 422 - mało znana czaszka odkryta w 1895 r. (historię opisują Brown i in. w książce z 2004 r. zatytułowanej "A Triceratops hunt in pioneer Wyoming") (Miller, online 2013). Długi, skierowany ku przodowi róg nosowy wskazuje, że to T. prorsus, nie jest jednak pewne, czy i na ile okaz jest zrekonstruowany. Ilustracje: Miller, online 2013. Lokalizacja: Wyoming (Miller, online 2013);
- $ MOR 1604 - niekompletna czaszka o długości całkowitej ok. 2 m i podstawowej 1,1 m, zawierająca przednią, niediagnostyczną część kryzy i zaliczona do stadium młodego dorosłego przez Scannellę i Hornera (2010) (Horner i Goodwin, 2006; 2008; Scannella i Horner, 2010; Scannella i in., 2014). Skierowany pionowo wyrostek nosowy kości przedszczękowej, długi (26,5 cm) róg nosowy i zredukowany (lub brak) guz na górnej powierzchni kości nosowej (za rogiem) wskazują, że to T. prorsus (Scannella i in., 2014). Ilustracje: MOR, online 2018; online 2020. Lokalizacja: górne środkowe / górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- ^ NDHC, bez nr – przypisany przez Farke (1997) do T. prorsus. Płaska powierzchnia kości łuskowej wskazuje, że to Triceratops a długi, skierowany ku przodowi róg nosowy, że T. prorsus. Prawdopodobnie jest to jeden z okazów z NDGS, wzmiankowanych niżej. Ilustracje: Hoganson, online 2006; North Dakota Geological Survey, online. Lokalizacja: formacja Hell Creek, Dakota Północna (Farke, 1997; Hoganson, online 2006).
(4) Materiał przypisany na podstawie wieku geologicznego (zob. #Legenda):
- ^ ? MOR 652 - zawiera kość łuskową młodego osobnika (Horner i Goodwin, 2006; Scannella i Fowler, 2014), która jest krótka, co sugeruje Triceratops. Ilustracje: Tanke, online 2014. Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- ^ $ MOR 2576 - izolowany róg nosowy o kształcie sugerującym T. ?sp. nov. lub T. prorsus (Scannella i in., 2015, ryc. 7.7A), lecz słabo wydłużonym (Scannella i in., 2014). Lokalizacja: górne górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- $ MOR 2979 - mniejsza część kryzy, rogi nadoczne, region ciemiączek (są zamknięte - Scannella i in., 2014) i k. przedszczękowa (Scannella i Fowler, 2014). Szeroka tylnobrzuszna powierzchnia dolnotylnego wyrostka kości przedszczękowej z wyraźnym grzbietem na bocznej powierzchni wskazuje, że to zaawansowany T. ?sp. nov. lub T. prorsus (Scannella i in., 2014). Ilustracje: Scannella i Fowler, 2014 , ryc. 13B. Lokalizacja: dolne górne osady formacji Hell Creek, hrabstwo Garfield, Montana (Scannella i Fowler, 2014; Scannella i in., 2014);
- ? MOR 3000 - m.in. częściowa kość ciemieniowa, kość łuskowa i róg nadoczny (Scannella i Fowler, 2014), kryza wydaje się raczej gruba, co sugeruje Triceratops [Boatman, 2012, ryc. 4.1]. Lokalizacja: dolne górne osady formacji Hell Creek, hrabstwo Garfield, Montana (Scannella i Fowler, 2014);
- ^ $ MOR 3049 - róg nosowy, którego długość (21 cm) i większy niż u T. horridus stopień wydłużenia wskazuje, że to T. ?sp. nov. lub T. prorsus (Scannella i in., 2014). Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014; Scannella i in., 2014);
- (?) UCMP 136306 - czaszka młodego osobnika o długości ok. 110 (Horner i Goodwin, 2006) lub 120 cm (Goodwin i Horner, 2014). Ma otwarte ciemiączko ciemieniowo-czołowe (ang. frontoparietal fontanelle - Farke, 2010), co sugerować może, że to T. horridus lub T. ?sp. nov., lecz wynika to prawdopodobnie z ontogenezy (Goodwin i Horner, 2014). Stosunek długości rogów nadocznych (38 i 40 cm - Goodwin i Horner, 2014) do długości od dzioba do kłykcia potylicznego (64 cm - Goodwin i Horner, 2014) mieści się w przedziale wszystkich gatunków, choć jest wysoki jak na T. prorsus (0,61). Kształt rogu nosowego sugeruje raczej T. ?sp. nov. lub T. prorsus (Scannella i in., 2015, ryc. 7.7A). Nie zachowały się m.in. kości przedszczękowe, szczękowe, nosowe. Czaszka opisana w: Goodwin i Horner, 2014. Ryan (2003, s. 268, podał numer 13306, który może odnosić się do tego okazu). Ilustracje: *; ** - #Media; Goodwin i Horner, 2014. Lokalizacja: górne osady formacji Hell Creek, hrabstwo McCone, Montana (Goodwin i Horner, 2014; Scannella i in., 2015, s. 426).
- ? UCMP 137263 - niekompletna czaszka o długości ok. 160 cm (Horner i Goodwin, 2006), zawiera większość kryzy (Goodwin i Horner, 2010, s. 558), której kształt określono jako „siodłowaty” („fan-shaped” - Horner i Goodwin, 2006, tab. 2), co sugeruje prawdopodobnie Triceratops. Kość łuskowa jest krótka, co również wskazuje na taką identyfikację (choć młode okazy Torosaurus również mogły mieć krótkie te kości). Ilustracje: Goodwin i Horner, 2010, ryc. 36.10. Lokalizacja: górne górne (najwyższe) osady formacji Hell Creek, hrabstwo McCone, Montana (Goodwin i Horner, 2010, s. 558; Scannella i Fowler, 2014; Scannella i in., 2014);
- ^ (?) UCMP 154452 - tylna część czaszki o dł. ok. 38 cm (do kłykcia ok. 28 cm [Goodwin i in., 2006], szacunki Scannelli i in. [2014] to 24,6 cm, 29,4 cm i 43,3 cm, pierwszy, jako oparty na długości żuchwy, jest zapewne najbliższy prawdy; szacunki Curriego i in. [2016] to 29 cm według długości żuchwy i 38 cm według kłykcia potylicznego) i żuchwa, fragmenty szkieletu pozaczaszkowego, opisane: w Goodwin i in., 2006. Ilustracje: *; ** - #Media; O; Goodwin i in., 2006. Lokalizacja: górne warstwy formacji Hell Creek, hrabstwo Garfield, Montana (Goodwin i in., 2006).
= łącznie 23 osobniki przypisane ponad wszelką możliwość oraz 30 mniej pewnych (=53), w tym 29 niewątpliwie należących do Triceratops.
Lokalizacje
W nawiasach kwadratowych podano liczbę okazów: USA - formacja Lance (Wyoming) [4], formacja Hell Creek (Montana [39], Dakota Południowa [3], Dakota Północna [2]); Kanada: formacja Frenchman (Saskatchewan) [2] i ??Scollard (Alberta, według Paul, 2010).
Etymologia
Prorsus oznacza "prosty" (ang. "straight on", Dodson, 1996), co odnosi się zapewne do skierowanego do przodu i tworzącego z dalszą powierzchnią czaszki niemal prostą linię, na co zwrócił uwagę Marsh (1890a).
Galeria
T. sp.

Warte uwagi okazy zaliczane do T. sp. to:
- AMNH 970 – duża czaszka Triceratops (zob. Torosaurus: Największe ceratopsy). Forster (1996b) i Anderson (1999) zaliczyli go do T. horridus a Longrich i Field (2012) oraz Longrich (2014) do T. prorsus. Otwarte ciemiączko czołowo-ciemieniowe (Forster, 1990, s. 55-56 i ryc. 2-7) u okazu w przedostatniej fazie rozwoju (Longrich i Field, 2012) może sugerować, że raczej nie jest to T. prorsus. Choć nie jest znane, w którym z etapów ontogenezy wyróżnionych przez Longricha i Fielda (2012) ciemiączko zarastało, wydaje się mało prawdopodobne, aby dopiero w ostatniej, skoro Torosaurus ANSP 15192, który również jest w przedostatniej fazie (Longrich i Field, 2012), ma je zamknięte (Farke, 2006). Ilustracje: *; Lull, 1903; Glut, 2002, s. 85. Lokalizacja: formacja Hell Creek, zachodnia Montana (Forster, 1996b);
- CNM 8862 (=GSC 8862) – holotyp T. albertensis - zob. niżej;
- okaz z DMC ("Larry") - prawie kompletny szkielet o długości ok. 7,6 m (=25 stóp) (DMC, online b). Ilustracje: DMC, online b;
- MCZ 1102 - holotyp T. eurycephalus - zob. niżej;
- MOR 1199 – czaszka młodego osobnika, której brakuje: rogu nosowego (Horner i Goodwin, 2006) oraz (contra Horner i Goodwin, 2006) kości rostralnej i przedzębowej (informacje z wystawy w MOR [Farke, online 2014b]) i prawdopodobnie innych elementów – np. górnych części rogów nadocznych i części kości przedszczękowej (Phillips, online 2016). Czaszka ma długość całkowitą 87 cm i podstawową 48,3 cm (Horner i Goodwin, 2006; Scannella i in., 2014). MOR 1199 został zaliczony do T. horridus przez Longricha i Fielda (2012) oraz Maiorino i in. (2013b). Analiza morfometryczna całej czaszki przez Maiorino i in. (2013b) wskazuje, że MOR 1199 należy do T. horridus, natomiast jej przód sugeruje T. prorsus. Ilustracje: *; O; Horner i Goodwin, 2006, ryc. 1b; Wolff, 2007, ryc. 5.1; Britannica, online 2010; Phillips online, 2016. Lokalizacja: ?środkowe osady formacji Hell Creek, hrabstwo McCone, Montana (Goodwin i Horner, 2010; Scannella i Fowler, 2014);
- ^ ? MOR 2946 - kość łuskowa i ciemieniowa z wklęsłym obszarem kryzy w miejscu, gdzie Torosaurus ma okna (Scannella i Horner, 2010) - nie jest jednak pewne, czy okaz nie miał małego okna w miejscu, które się nie zachowało. Ilustracje: Scannella i Horner, 2010, ryc. 1C; Fowler, online 2020. Lokalizacja: nieznana, formacja Hell Creek, Montana (Scannella i Fowler, 2014);
- NSM PV 20379 ("Raymond", WFQ:WRG 94.014) - artykułowany, prawie kompletny szkielet - bez większości lewej strony, ogona i przedniej części pyska (Fujiwara, 2009). Szacowana długość szkieletu bez czaszki to ok. 5,3 m a dodając do tego czaszkę (ok. 94 cm - Fujiwara, 2009, ryc. 2) całkowita długość to ok. 6,14 m. Garstka i Burnham (1997) oraz Russell i Manabe (2002) zaliczyli okaz do T. horridus; pierwsi autorzy wyróżniali tylko ten gatunek a drudzy prawdopodobnie jedynie powtórzyli taką klasyfikację. Według Fujiwary (2009) jest to Triceratops sp., zaś Longrich (2014) użył publikacji Fujiwary do kodowania w analizie T. prorsus. Wydaje się, że okaz ma mały róg nosowy, co sugerowałoby T. horridus, lecz z powodu niekompletności i możliwej deformacji (kość przedczołowa jest wysunięta nad oczodołem ku górze – ryc. 2 w Fujiwara, 2009) nie jest to pewne. Ilustracje: *; ** - #Diagnoza; Fujiwara, 2009, ryc. 2. Lokalizacja: formacja Hell Creek - 17 m poniżej kontaktu z formacją Fort Union (górne osady?), hrabstwo Bowman, Dakota Północna (Fujiwara, 2009);
- TLAM G.BA.2004.024.0001 - gruba na 5,2 cm kość łuskowa, poważnie uszkodzona za życia zwierzęcia (Farke i Alley, 2006). Ilustracje: Farke i Alley, 2006. Lokalizacja: formacja Hell Creek; hrabstwo Corson, Dakota Południowa (Farke i Alley, 2006);
- USNM 4842 - Hatcher i in. (1907, s. 128), Lull (1933, s. 119) oraz Ostrom i Wellnhofer (1986, s. 130) podają, że to szkielet pozaczaszkowy oraz niekompletny róg nadoczny (choć na ryc. 108 w tej pierwszej publikacji został zilustrowany w całości). Mimo niekompletności, okaz został zaliczony do T. prorsus przez Hatchera i in., Gilmore'a (1906); klasyfikację taką podają też Lyson i Longrich (2011) oraz Ford (online - Triceratops). W zmontowanym okazie umieszczono czaszkę USNM 2100 (Ostrom i Wellnhofer, 1986, s. 130). Okaz należy do Triceratops ze względu na budowę łopatki (zob. opis Torosaurus) (Gilmore, 1906; Milner, online 2014; okaz ma łopatkę - Hatcher i in., 1907, s. 128). Ilustracje: *; ** - #Media; Schuchert, 1905; Gilmore, 1906. Lokalizacja: formacja Lance (Lyson i Longrich, 2011), hrabstwo Converse (Niobrara?), Wyoming (Gilmore, 1906);
- UWGM 732 - przynajmniej kość łuskowa, ciemieniowa i rogi nadoczne. Okaz nie ma zgrubienia na kości łuskowej, należy więc do Triceratops (Scannella i Horner, 2010; contra Lyson i Longrich, 2011 oraz Brown i in., 2021, którzy zaliczyli go do Torosaurus latus). Kość łuskowa mierzy ok. 1 m, co jest obok MOR 2702 wartością najwyższą wśród Triceratops; jest też jedną najbardziej wydłużonych (Scannella i Horner, 2010, ryc. 4). Zapewne jest to kość łuskowa wzmiankowana przez Penkalskiego i Skulana (2010) (Keenan i Scannella, 2014, s. 359). Ilustracje: Scannella i Horner, 2010, ryc. 3I. Lokalizacja: formacja Hell Creek, hrabstwo Carter, Montana (Penkalski i Skulan, 2010; Lyson i Longrich, 2011).
Inne okazy, które można zaliczyć do Triceratops to:
- ^ $ AMNH 972 - przypisany do Triceratops (Forster, 1990; 1996a, s. 248) róg nosowy i kość przedzębowa (AMNH, online – 972). Róg jest znacznie wydłużony, co wskazuje na zaawansowanego T. ?sp. nov. lub T. prorsus. Ilustracje: Forster, 1990, ryc. 1-5;
- BYU 19974 – duży okaz, który ma cienką kość ciemieniową w miejscu okien Torosaurus (Beach, 2011). Brak okien sugeruje przynależność do Triceratops;
- CMN 8741 – praktycznie kompletna kryza, okolica oczodołu i kości jarzmowej (Tanke, online 2018). Kość łuskowa według badań morfometrycznych należy do Triceratops (Maiorino i in., 2013b). Według suplementu tej pracy jest to T. sp. a według ryciny 6 to T. horridus. Z uwagi na wiek formacji Frenchman, korelujący z górnymi i częścią środkowych warstw formacji Hell Creek, mało prawdopodobne jest występowanie w niej T. horridus (Scannella i in., 2014; Eberth i Kamo, 2020). Ilustracje: Tanke, online 2018. Lokalizacja: formacja Frenchman, Grasslands National Park, Saskatchewan (Tokaryk, 2009);
- DMNH 17060 – wielki (zob. Torosaurus: Największe ceratopsy) częściowy szkielet niezawierający czaszki (żuchwa, łopatka, kość krucza, kilka kręgów, kość ramienna, miednica, fragmenty kończyny tylnej), zidentyfikowany jako cf. Torosaurus? sp. na podstawie budowy kości kruczej i ramiennej (Carpenter i Young, 2002), które jednak nie pozwalają na rozróżnianie Torosaurus i Triceratops, lecz pozwala na to budowa łopatki, która u tego okazu ukazuje budowę diagnostyczną dla Triceratops (zob. opis Torosaurus). Lyson i Longrich (2011) oznaczyli go jako ?Torosaurus sp. Ilustracje: Carpenter i Young, 2002, ryc. 11. Lokalizacja: formacja Laramie (Lance?), Leyden Gulch, Colorado (Carpenter i Young, 2002; Lyson i Longrich, 2011).
- ^ LACM 149538 - kość łuskowa młodego osobnika o długości 33 cm (szacowana długość czaszki 68 cm) (Hurt, online). Brak zgrubienia wzdłuż wewnętrznej krawędzi i kształt (Maiorino i in., 2013b) sugeruje Triceratops. Według suplementu Maiorino i in. (2013b) jest to T. sp., lecz według ryciny 6 to niepewny T. horridus. Ilustracje: Hurt, online; LACM, online. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Maiorino i in., 2013b; LACM, online);
- ? ^ LACM 154911 - kość łuskowa młodego osobnika, według badań morfometrycznych należąca do Triceratops (Maiorino i in., 2013b). Według suplementu tej publikacji jest to T. sp., lecz według ryciny 6 to niepewny T. horridus. Według informacji z bazy okazów LACM jest to odlew (LACM, online). Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (Maiorino i in., 2013b; LACM, online);
- $ MOR 965 - został zaliczony do Triceratops (Tanke i Farke, 2006, s. 328), ale też do Torosaurus (MOR, online). Zawiera m.in. znacznie dłuższy (24,9 cm) niż u wszystkich Torosaurus i T. horridus, lecz niewydłużony (szerokości 17 cm) róg nosowy, części kryzy (Scannella i Fowler, 2014; Scannella i in., 2014) oraz kość jarzmową i kwadratową (Tanke i Farke, 2006, s. 328 i s. 346). Tak duże rozmiary rogu spotykane są jedynie u Triceratops, lecz są one znacznie węższe (znanych jest kilka rogów o takich proporcjach – zob. Komentarz do cech w ewolucji Triceratops: cecha 6). Lokalizacja: formacja Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 2551 - zawiera kryzę (MOR, online) - kości ciemieniowe (Farke i in., 2009), róg nadoczny (Scannella i Horner, 2018) oraz róg nosowy o długości 22 cm (Scannella i in., 2014), co sugeruje, że to Triceratops. Scannella i Horner (2018) określają go jako "large non-toromorph", co potwierdza, że ma morfologię Triceratops. Lokalizacja: górne/środkowe osady formacji Hell Creek, Montana (Horner i in., 2011; Scannella i Fowler, 2014);
- ^ MOR 2572 - kość łuskowa młodego osobnika, nie ma zgrubienia, więc należy do Triceratops [Goodwin i Horner, 2010, ryc. 13.3]. Lokalizacja: formacja Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 2969 - kość ciemieniowa długości ok. 60 cm (Horner i Lamm, 2011), której grubość (ponad 6 cm; na ryc. 2A w Horner i Lamm, 2011 ma ok. 4,4 cm) wskazuje, że jest to Triceratops (Scannella i Horner, 2010). Lokalizacja: formacja Hell Creek, Montana (Scannella i Fowler, 2014);
- NHMUK R3886-8 (lub R3886; również jako NHMUK PV R 3886) – niekompletna czaszka i szkielet pozaczaszkowy, zaliczone do T. prorsus (Maidment i Barrett, 2011). Budowa kości łuskowej wskazuje na Triceratops, lecz przednia część czaszki jest niekompletna, choć róg nosowy wydaje się mały i wąski – może być niekompletny (Graham, online 2016) [Graham, online 2016]. Lokalizacja: formacja Lance/Hell Creek, Wyoming (Milner i Barrett, 2016);
- RSM P2982.1 - kompletna czaszka niedorosłego osobnika (ang. subadult sensu Horner i in., 2006) z rogiem nadocznym mierzącym 58,5 cm (Bamforth, 2018). Prawdopodobnie jest to okaz zilustrowany przez Bamforth (online 2019a - por. Bamforth, 2018 oraz Tokaryk, 2009). Ze zdjęcia wynika, że kryza jest gruba, nie ma okien ani zgrubienia na kości łuskowej zaś róg nosowy wydaje się raczej wydatny, co wskazuje na zaawansowanego Triceratops. Ilustracje: ? Bamforth, online 2019a. Lokalizacja: dolina rzeki Frenchman, okolica Eastend, Chambery Coulee (Bamforth, 2018; Tokaryk, 2009);
- okaz z SMF (okaz 1 - z kryzą) – tylna część czaszki (Drevermann, 1913; contra SMF, online 2011 – zob. zdjęcie w SMF, online 2011), która ma otwarte ciemiączko czołowo-ciemieniowe [*; Drevermann, 1913; SMF, online 2011]. Lokalizacja: formacja Lance, Wyoming (SMF, online 2011);
- ? okaz z SMF (okaz 2 - bez kryzy) - czaszka niezawierająca kryzy, lecz zaliczana do T. prorsus (Drevermann, 1913; SMF, online 2011). Wielkość i kształt rogu nosowego raczej wyklucza T. horridus i Torosaurus. Ilustracje: *; SMF, online 2011. Lokalizacja: formacja Lance, Wyoming (SMF, online 2011);
- ^ $ TMP 1984.039.0012 - zaliczony do Triceratops (Tanke i Farke, 2006), zawiera przynajmniej róg nosowy, którego wydłużony kształt wyklucza T. horridus i Torosaurus. Ilustracje: Tanke i Farke, 2006, ryc. 14.9. Lokalizacja: formacja Lance, Wyoming (TMP, online);
- ? UCMP 129205 - elementy kryzy, której kształt określono jako „siodłowaty” („fan-shaped” - Horner i Goodwin, 2006, tab. 2), co sugeruje prawdopodobnie Triceratops. Według bazy danych UCMP (online) to kości nosowe i przedszczękowa. Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (UCMP, online);
- USNM 5741 - Farke (online 2012a) podaje, że to Triceratops z powodu grubości kości łuskowej (kształt Farke określił jako "squared off" - jak u Ojoceratops). Wcześniej okaz do Triceratops zaliczyła Forster (1990). Na zdjęciu (USNM, online) widoczny jest m.in. róg o szacowanej długości min. 40 cm oraz pokryte siatką naczyń fragmenty kryzy. Ilustracje: USNM, online. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (USNM, online);
- ^ USNM 6679 - tylna część czaszki (Gilmore, 1920, s. 104), która nie ma okien w kryzie. Ilustracje: Gilmore, 1920, tabl. 7. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Lull, 1915);
- YPM 1821 - holotyp T. flabellatus - zob. niżej.
- YPM 1823 - holotyp T. serratus - zob. niżej.
= łącznie 20 osobników przypisanych ponad wszelką możliwość oraz 8 mniej pewnych (=26).
= łącznie 90 niewątpliwych osobników przypisanych do rodzaju (razem z okazami zaliczonymi do gatunków), z mniej pewnymi 138 (w tym 21 okazów zaliczonych ze względu na budowę zaawansowanych Triceratops).
?T. sp.
Poniżej wyliczono materiał zaliczany do Triceratops, lecz z różnych powodów nie ma pewności, czy do niego rzeczywiście należy. Materiał pogrupowano według danych mających wskazywać na przynależność do Triceratops, ułożone według stopnia pewności przynależności (zdjęcia okazów obok to okazy pewne, należące do T. horridus). Nie oznaczano okazów mogących należeć do [[Tatankaceratops.
Materiał zaliczany do Triceratops i zawierający części kryzy - prawdopodobnie większość jest diagnostyczna i należy rzeczywiście do Triceratops :
- ASU bez nr, zawiera kość łuskową (Farke i in., 2009), która wygląda na płaską, krótką i z zakrzywioną boczną krawędzią, co sugeruje Triceratops (ASU News, online 2009);
- CMN 8861 - zaliczony do Triceratops (Eberth, 1997; Eberth i in., 2013), zawiera kość łuskową (Farke, online 2013). Lokalizacja: górne warstwy formacji Scollard, Ardley, Alberta (Eberth, 1997; CMN, online);
- CMN 9542 - zaliczony do Triceratops (Russell i Singh, 1978; Lerbekmo i in., 1979; Eberth, 1997; Lyson i Longrich, 2011; Eberth i in., 2013), zob. też opis Torosaurus - okaz UALVP 1646. Lokalizacja: górne warstwy formacji Scollard, Alberta (Eberth, 1997; Eberth i in., 2013);
- CNM 34824 - prawie kompletna czaszka (Tokaryk, 2009), zawiera kryzę (Barrick i in., 1998). Lokalizacja: formacja Frenchman, Saskatchewan (Tokaryk, 2009; Lyson i Longrich, 2011; CNM, online);
- CMN 56508 - kryza i rogi nadoczne (McDonald, online 2018a), może zawierać odcisk skóry z kryzy (McDonald, online 2018b; Bell i in., 2022). Ilustracje: McDonald, online 2018a; online 2018b. Lokalizacja: formacja Frenchman, Eastend, Saskatchewan (McDonald, online 2018a; Bell i in., 2022);
- CSC-4-I – zawiera rogi i kryzę (Farke, 1997), na zdjęciu widoczne jest, że znaczna część kryzy jest zrekonstruowana. Ilustracje: ?Swanson, online 2013 – por. CSC, online. Lokalizacja: formacja Lance (Farke, 1997);
- LACM 151465 – kość łuskowa (LACM, online). Lokalizacja: formacja Hell Creek, hrabstwo Carter, Montana (LACM, online);
- LACM 151472 – fragmenty kryzy (LACM, online). Ilustracje: LACM, online. Lokalizacja: formacja Hell Creek, hrabstwo Carter, Montana (LACM, online);
- MOR 2950 - zawiera m.in. większość kości łuskowej i część ciemieniowej (Scannella i Fowler, 2014), zaliczone według kryteriów Scannelli i Hornera do stadium młodocianego - subadult (Scannella i Horner, 2010). Lokalizacja: górne górne (najwyższe) osady formacji Hell Creek, Montana (Scannella i Fowler, 2014; Scannella i in., 2014);
- PTRM V88005 (nazwa lokalizacji) - zawiera fragmenty kryzy, zaliczony do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: górne osady formacji Hell Creek, hrabstwo Bowman, Dakota Północna (Pearson i in., 2002);
- PTRM V88040 (nazwa lokalizacji) - kryza, zaliczona do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: górne osady formacji Hell Creek, hrabstwo Bowman, Dakota Północna (Pearson i in., 2002);
- PTRM V91002 (nazwa lokalizacji) - zawiera m.in. kość łuskową, według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: dolne osady formacji Hell Creek, hrabstwo Bowman, Dakota Północna (Pearson i in., 2002);
- PTRM V91012 (nazwa lokalizacji) - zawiera fragmenty kryzy, zaliczony do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: dolne osady formacji Hell Creek, Dakota Północna (Pearson i in., 2002; Lyson i Longrich, 2011);
- PTRM V95009 (nazwa lokalizacji) - zawiera fragmenty kryzy, zaliczony do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: górne osady formacji Hell Creek, hrabstwo Slope, Dakota Północna (Pearson i in., 2002);
- PTRM V95014 (nazwa lokalizacji) - zawiera kryzę (Pearson i in., 2002), zaliczony do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: górne osady formacji Hell Creek, hrabstwo Slope, Dakota Północna (Pearson i in., 2002);
- PTRM V95017 (nazwa lokalizacji) - fragmenty kryzy, zaliczony do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: górne osady formacji Hell Creek, hrabstwo Slope, Dakota Północna (Pearson i in., 2002);
- PTRM V95023 (nazwa lokalizacji) - zawiera fragmenty kryzy (Pearson i in., 2002), zaliczony do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: górne osady formacji Hell Creek, hrabstwo Slope, Dakota Północna (Pearson i in., 2002);
- ? PTRM V95104 (nazwa lokalizacji) - zawiera kryzę, zaliczony do Triceratops (Lyson i Longrich, 2011). Autorzy ci cytują publikację Pearsona i in. (2002), lecz nie ma w niej informacji o takim numerze; nie jest to też błąd w numerze V95014, skoro jest on osobno w suplemencie pracy Lysona i Longricha (2011). Lokalizacja: formacja Hell Creek, Dakota Północna (Lyson i Longrich, 2011);
- PTRM V96019 (nazwa lokalizacji) - zawiera kryzę, zaliczony do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: górne osady formacji Hell Creek, hrabstwo Slope, Dakota Północna (Pearson i in., 2002);
- PTRM V96021 (nazwa lokalizacji) - zawiera kryzę, zaliczony do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: górne osady formacji Hell Creek, hrabstwo Slope, Dakota Północna (Pearson i in., 2002);
- PTRM V97019 (nazwa lokalizacji?) - zawiera kryzę, zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Dakota Północna (Lyson i Longrich, 2011);
- PTRM V98015 (nazwa lokalizacji) - kryza, zaliczona do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: górne osady formacji Hell Creek, hrabstwo Slope, Dakota Północna (Pearson i in., 2002);
- PTRM V98016 (nazwa lokalizacji) - kryza (Lyson i Longrich, 2011) lub kość ciemieniowa (Pearson i in., 2002), zaliczona do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: górne osady formacji Hell Creek, Dakota Północna (Pearson i in., 2002);
- SMM P60/2/1 (zapisywany również P60.2.1) - zawiera niekompletną czaszkę (Erickson, 1996), w tym kość łuskową, zaliczoną do stadium młodego dorosłego Triceratops przez Scannellę i Hornera (2010). Okaz został przez Ericksona (1996) zaliczony do T. prorsus a róg nosowy ma kształt sugerujący ten gatunek albo T. ?sp. nov. (Scannella i in., 2015, ryc. 7A), lecz z uwagi na obecność w tym badaniu tylko jednego z okazów z SMM, może to dotyczyć SMM P62/1/1. Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (Erickson, 1966);
- TLAM bez nr - zawiera kości łuskowe i ciemieniową (Farke i in., 2009);
- TMP 1998.102.0001 – zęby i izolowane elementy zaliczone do Triceratops (Ryan, 2003, s. 565; Eberth i in., 2013). Lokalizacja: dolne warstwy formacji Scollard, Alberta (Eberth i in., 2013);
- okaz z nieznanej instytucji (bez nr, Eberth i in., 2013, tabela A1, czwarty wpis o Triceratops) - zaliczony do Triceratops (Eberth i in., 2013). Lokalizacja: dolne warstwy formacji Scollard, Alberta (Eberth i in., 2013);
- TMP 2009.125.0001, zaliczony ze znakiem zapytania do Triceratops (Eberth i in., 2013). Lokalizacja: dolne warstwy formacji Scollard, Alberta (Eberth i in., 2013);
- UCM 13745 - fragment kryzy, zaliczony do Triceratops (Carpenter, 1979). Lokalizacja: formacja Laramie, hrabstwo Weld, Colorado (Carpenter, 1979);
- UCMP 136589 - zawiera fragment kości łuskowej (Horner i Goodwin, 2006; Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, hrabstwo McCone, Montana (Lyson i Longrich, 2011; UCMP, online);
- UCMP 150234 - kość ciemieniowa młodego osobnika (Horner i Goodwin, 2006). Według bazy danych UCMP (online) jest to krokodylomorf;
- UCMP 150324 - proksymalny fragment kości ciemieniowej (UCMP, online). Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (UCMP, online);
- UCMP 159233 - m.in. fragmenty kryzy i róg nadoczny (Scannella i Horner, 2010; Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (Lyson i Longrich, 2011; UCMP, online);
- UCMP 172363 - zawiera fragment(y?) kryzy (UCMP, online). Lokalizacja: formacja Hell Creek, hrabstwo Slope, Montana (UCMP, online);
- UCMP 174838 - zawiera element kryzy (Horner i Goodwin, 2006), określony jako "co najmniej dorosły - nie w pełni wyrośnięty" i przebadany histologicznie - szew kości jarzmowej i rogu (Bailleul i Horner, 2016). Róg nosowy jest "znacznie rozrośnięty" (Goodwin i Horner, 2001). Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (Lyson i Longrich, 2011; Bailleul i Horner, 2016; UCMP, online);
- UCMP 91MG-7-11-4 - zawiera kość łuskową (Farke i in., 2009). Nie ma takiego okazu według bazy danych UCMP (online);
- USNM 2124 - zaliczany do Triceratops (Lull, 1915, s. 343; 1933, s. 15; Butler i Barrett, 2008; Lyson i Longrich, 2011), według Farke i in. (2009) oraz Lysona i Longricha (2011) zawiera kość łuskową. Ilustracje: USNM, online. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (USNM, online);
- USNM 2416 - zaliczany do Triceratops (Lull, 1915, s. 343; 1933, s. 15; Gilmore, 1920; Ostrom i Wellnhofer, 1986, s. 117; Forster, 1990; 1996a; Butler i Barrett, 2008; Lyson i Longrich, 2011; Farke, online 2012b; USNM, online; Borkovic, 2013), zawiera kość ciemieniową (Farke, online 2012b) i kości łuskowe (Farke i in., 2009). Borkovic (2013) zaliczył go do T. horridus. Longrich (2011) podaje, że to Pentaceratops. Ilustracje: Gilmore, 1920, ryc. 2-3; USNM, online. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (USNM, online);
- USNM 2417 - zawiera fragmenty kryzy, zaliczone do Triceratops (Lyson i Longrich, 2011). Ilustracje: USNM, online. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (USNM, online);
- USNM 4286 - zaliczany do Triceratops (Hatcher i in., 1907, s. 134; Lull, 1915, s. 343; 1933, s. 15; Ostrom i Wellnhofer, 1986; Forster, 1990; 1996a; Holliday, 2009; Lyson i Longrich, 2011, Wolff, 2007; Borkovic, 2013), według Lysona i Longricha (2011) oraz Farke i in. (2009) zawiera kość łuskową (Farke i in., 2009), więc powinien być diagnostyczny, jednak został zaliczony tylko do Ceratopsidae przez Butlera i Barretta (2008). Okaz ma mały i spiczasty róg nosowy (Lull, 1933, s. 129) oraz głęboką bruzdę na przedniej powierzchni rogów nadocznych (Hatcher i in., 1907, s. 134; Lull, 1933, s. 129). Ilustracje: Hatcher i in., 1907, ryc. 113. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Lull, 1933);
- ? USNM 4741 - zaliczony do Triceratops (Longrich i Field, 2012), zawiera kość łuskową (Longrich i Field, 2012) i ciemieniową (Maiorino i in., 2013b), według USNM (online) jest to Uintatherium; żadna z publikacji nie podaje naraz USNM 4741 i 5741, co sugeruje, że oznaczenie może być pomyłką pisarską;
- USNM 5738 - zaliczany do Triceratops (Lull, 1915, s. 343; 1933, s. 15; Butler i Barrett, 2008; Lyson i Longrich, 2011), według Farke i in. (2009) oraz Lysona i Longricha (2011) zawiera kość łuskową. Ilustracje: USNM, online. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (USNM, online);
- USNM 5740 - T. sp. (Lull, 1933, s. 15, Forster, 1990; 1996a; Holliday, 2009; Longrich i Field, 2012; Farke, online 2012b; Paulina-Carabajal i in., 2016), zawiera kość łuskową (Longrich i Field, 2012; Farke, online 2013b). Prawa część okazu nosi oznaczenie CNM 8528 (Forster, 1996a, s. 250; Farke, online 2013) i jest zaliczana do Triceratops (Forster, 1996a; Anderson 1999; Farke, 2010; Holliday, 2009). Ilustracje: Gilmore, 1920, ryc. 1; Gilmore, 1920, tabl. 8.Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Lull, 1933);
- USNM 5786 - zaliczany do Triceratops (Lyson i Longrich, 2011), zawiera kość łuskową (Farke i in., 2009; Lyson i Longrich, 2011; USNM, online). Ilustracje: USNM, online. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (USNM, online);
- USNM 6525 - zawiera kość łuskową (USNM, online) - jej przednio-boczną część (Farke, online 2012b) a według Farke i in. (2009) także ciemieniową. Ilustracje: USNM, online. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (USNM, online);
- USNM 7239 - zaliczany do Triceratops (Lull, 1915, s. 343; 1933, s. 15), zawiera kość łuskową (USNM, online; Farke i in., 2009). Ilustracje: USNM, online. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (USNM, online);
- USNM 8081 - zaliczany do Triceratops (Lyson i Longrich, 2011; Wolff, 2007; Maiorino i in., 2017), zawiera fragmenty kryzy (Lyson i Longrich, 2011)/kości łuskowej (USNM, online) i dobrze zachowaną kość przedszczękową i dziób (Farke, online 2012b). Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Farke, online 2012; USNM, online);
- USNM "Skull 15" - zawiera kość łuskową (Farke i in., 2009);
- ?? YPM 1824 - zawiera kość łuskową (Wu i in., 2007), według YPM (online) jest to Knightia. Brak odesłań w literaturze do tego okazu i podobieństwo do numerów innych okazów z YPM sugeruje, że jest to pomyłka pisarska;
- ?? YPM 2046 - zawiera kość łuskową (Farke i in., 2009); okaz o takim numerze to odcisk stopy, holotyp ichnotaksonu Otozoum minus (Rainforth, 2003; YPM, online).
- YPM VPPU 018159 - m.in. część kryzy (YPM, online). Lokalizacja: formacja Lance, hrabstwo Park, Wyoming (YPM, online);
- YPM VP 053263 - m.in. kość łuskowa ze śladami ugryzienia. Ilustracje: Longrich i in., 2010, ryc. 1D. Lokalizacja: formacja Lance, Wyoming (YPM, online).
= 50 okazów.
Okazy przypisane przez Farkego (2006), wyraźnie rozróżniającego Triceratops i Torosaurus, lecz przypisanie izolowanego rogu nadocznego MOR 539 podważa przypisania:
- GP 246-A - min. róg nadoczny (Farke, 2006);
- MOR 539 - róg nadoczny, prawdopodobnie wyłącznie (Forster, 1996a, s. 249; Farke, 2006; Horner i Goodwin, 2006; Scannella i Fowler, 2014). Lokalizacja: formacja Hell Creek, Montana (Scannella i Fowler, 2014).
- SDSM 3110 - min. róg nadoczny (Farke, 2006);
- SDSM 58721 - min. róg nadoczny (Farke, 2006);
= 4 okazy.
Okazy o nieznanym materiale, zaliczone przez Lysona i Longricha (2011); autorzy ci zaliczali do Triceratops także ewidentnie niediagnostyczne okazy (np. kręgi PTRM v97009):
- GCM nieskatalogowany 1 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 2 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 3 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 4 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 5 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 6 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 7 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 8 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 9 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 10 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 11 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 12 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 13 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 14 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 15 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 16 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- GCM nieskatalogowany 17 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- LACM 152525 - zawiera czaszkę zaliczoną do Triceratops (Lyson i Longrich, 2011), lecz w bazie LACM podano, że to szkielet pozaczaszkowy (LACM, online). Lokalizacja: formacja Hell Creek, hrabstwo Carter, Montana (Lyson i Longrich, 2011; LACM, online);
- LACM 7245 - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- MRF v01NOR - zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Dakota Północna (Lyson i Longrich, 2011);
- MRF v03TRI2 - zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Dakota Północna (Lyson i Longrich, 2011);
- MRF v05JP - zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Dakota Północna (Lyson i Longrich, 2011);
- MRF v06HOM - zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Dakota Północna (Lyson i Longrich, 2011);
- MRF v06JUV - zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Dakota Północna (Lyson i Longrich, 2011);
- MRF v06NOR - zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Dakota Północna (Lyson i Longrich, 2011);
- MRF v06VIR - zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Dakota Północna (Lyson i Longrich, 2011);
- MRF v07TS - zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Dakota Północna (Lyson i Longrich, 2011);
- MRF v08BB - zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Dakota Północna (Lyson i Longrich, 2011);
- MRF v09PP - zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- NDGS 06-3.1 - zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Dakota Północna (Lyson i Longrich, 2011);
- NDGS 555 - zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Dakota Północna (Lyson i Longrich, 2011);
- PTRM V88019 - czaszka zaliczona do Triceratops (Lyson i Longrich, 2011). Lokalizacja: górne osady formacji Hell Creek, hrabstwo Slope, Dakota Północna (Pearson i in., 2002);
- PTRM V92066 - zaliczony do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: środkowe osady formacji Hell Creek, hrabstwo Bowman, Dakota Północna (Pearson i in., 2002);
- PTRM V93005 - zaliczony do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: górne/środkowe osady formacji Hell Creek, hrabstwo Bowman, Dakota Północna (Pearson i in., 2002);
- PTRM V96016 - zaliczony do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: górne osady formacji Hell Creek, hrabstwo Bowman, Dakota Północna (Pearson i in., 2002);
- PTRM V97017 - zaliczony do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: górne osady formacji Hell Creek, hrabstwo Slope, Dakota Północna (Pearson i in., 2002);
- RSM 2525.1 - częściowa czaszka zaliczona do Triceratops (Lyson i Longrich, 2011), zawierająca kryzę i rogi nadoczne (Tokaryk, 2009) o długości 85 cm (Bamforth, 2018). Zapewne to ten samo okaz co SMNH P2525.1 (Farke, 2006). Lokalizacja: formacja Frenchman, Saskatchewan (Lyson i Longrich, 2011; Bamforth, 2018);
- RSM P2642 - częściowa czaszka zaliczona do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Frenchman, Saskatchewan (Lyson i Longrich, 2011);
- RSM nieskatalogowany 1 - czaszka i kończyny zaliczone do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Frenchman, Saskatchewan (Lyson i Longrich, 2011);
- RSM nieskatalogowany 2 - częściowa czaszka zaliczona do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Frenchman, Saskatchewan (Lyson i Longrich, 2011);
- UCMP 139292 - fragmenty czaszki zaliczone do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (Lyson i Longrich, 2011; UCMP, online);
- TPI: TTFKA-Val (instytucja nieznana) - szkielet zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Dakota Południowa (Lyson i Longrich, 2011).
= 42 okazy.
Wybrane okazy z kolekcji, w których do Triceratops zalicza się również ewidentnie niediagnostyczne okazy: w CM np. żebra (CM 83086), kręgi szyjne (CM 247), paliczki (CM 255) (Henrici, 2016):
- CM 1618 - zawierający m.in. kość dziobową, łuskową, krzyżową, łopatkę (106 cm), kość ramienną (68-69,6 cm), kości miednicy, kość udową (126,9 cm) i piszczel okaz zaliczany do Triceratops (Henrici, 2016; McIntosh, 1981, s. 42; Chinnery, 2004; Maidment i in., 2012). Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Henrici, 2016; McIntosh, 1981, s. 42);
= 1 okaz (wybrano nieliczne, warte odnotowania okazy).
Okazy z MOR, zaliczane do Triceratops przez autorów wyróżniających tylko ten rodzaj ceratopsyda z późnego mastrychtu - począwszy od publikacji Scannelli i Hornera (2010). Jedynie dwa z poniższych okazów (MOR 965 i MOR 2570) zostały zaliczone przed tym badaniem. Okazy te zawierają diagnostyczne elementy kryzy (kości łuskowe lub ciemieniowe), lecz niektóre mogą należeć innych ceratopsydów, zwłaszcza Torosaurus, gdyż zaliczane przez tych autorów do Triceratops okazy wykazują, jak wynika z ich późniejszych publikacji, budowę Torosaurus. Dotyczy to MOR 2984 (Triceratops według: Scannella i Horner, 2010; Horner i in., 2011; Scannella i Fowler, 2014), który wykazuje budowę Torosaurus (Scannella i in. 2015, s. 448) i MOR 3005 (Triceratops według Scannella i Fowler, 2014), który ma cienką kryzę, charakterystyczną dla Torosaurus (Scannella i in. 2014; zob. szerzej w Torosaurus: Materiał kopalny). MOR 2984 i MOR 3005 zostały wykreślone z niniejszej listy po ujawnieniu, że są to prawdopodobnie Torosaurus:
- MOR 335 - zawiera kość łuskową (MOR, online) lub ciemieniową (Farke i in., 2009; Scannella i Horner, 2010; Scannella i Fowler, 2014). Lokalizacja: dolne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 1053b - zawiera kość łuskową (Farke i in., 2009);
- MOR 1186 - zawiera m.in fragmenty kryzy (MOR, online) / kość ciemieniową (Farke i in., 2009; Scannella i Fowler, 2014), kość przedszczękową (Scannella, 2020) i elementy pozaczaszkowe (Scannella i Fowler, 2014). Okaz ma wymiary porównywalne z MOR 1122 7-22-00-1 (Scannella, 2020). Lokalizacja: dolne-środkowe osady formacji Hell Creek, Montana (Scannella i Fowler, 2014; dolne podają Scannella i in., 2015, s. 448; dolną połowę podał Scannella, 2020);
- MOR 2570 - zawiera m.in. fragmenty kości łuskowe (Horner i Goodwin, 2008; Scannella i Fowler, 2014), określony jako młody dorosły i przebadany histologicznie - szew kości jarzmowej i rogu (Bailleul i Horner, 2016). Lokalizacja: dolne środkowe osady formacji Hell Creek, hrabstwo Garfield, Montana (Scannella i Fowler, 2014; Scannella i in., 2014);
- MOR 2937- zawiera m.in. fragment kryzy (Scannella i Fowler, 2014) albo kość szczękowa (Scannella i Horner, 2010). Lokalizacja: górne górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 2597 - m.in. fragment kryzy (Goodwin i Horner, 2010; Scannella i Fowler, 2014). Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 2927 - zawiera kość łuskową (Scannella i Horner, 2010; Scannella i Fowler, 2014) lub ciemieniową (Farke i in., 2009). Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 2928 - zawiera kość łuskową (Scannella i Horner, 2010). Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 2929 - kość łuskowa młodego osobnika (Scannella i Horner, 2010; Scannella i Fowler, 2014). Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 2942 - kość łuskowa (Scannella i Horner, 2010; Scannella i Fowler, 2014). Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 2959 - zawiera m.in. fragmenty kryzy (Scannella i Fowler, 2014). Lokalizacja: ?dolne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 2970 - kość łuskowa (Scannella i Horner, 2010);
- MOR 2975 - m.in. kość łuskowa (Scannella i Horner, 2011; Scannella i Fowler, 2014). Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 2980 - k. łuskowa (Scannella i Horner, 2010; Scannella i Fowler, 2014). Lokalizacja: dolne górne osady formacji Hell Creek, hrabstwo Garfield, Montana (Scannella i Fowler, 2014);
- MOR 2982 - m.in. częściowa kość łuskowa, środkowa część k. ciemieniowej, dziób, kość i róg nosowy, rogi nadoczne, kość jarzmowa (Scannella i Fowler, 2014). Okaz jest w analizach bardziej zaawansowany niż T. horridus MOR 1120 i ma budowę zbliżającą go do zaawansowanego Triceratops ?sp. nov. (Scannella i Fowler, 2014), co może wykluczać, że to Torosaurus, lecz nieco bardziej zaawansowaną budowę rogu nosowego ma Torosaurus YPM 1831 (Scannella i in., 2015, ryc. 7.7.A). Ilustracje: Scannella i in., 2014, ryc. 1F. Lokalizacja: dolne środkowe osady formacji Hell Creek, hrabstwo Garfield, Montana (Scannella i Fowler, 2014);
- MOR 2991 - zawiera część kryzy. Lokalizacja: formacja Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 2992 - część kości łuskowej. Lokalizacja: formacja Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 3004 - kompletna duża czaszka i elementy pozaczaszkowe (Scannella i Fowler, 2014). Lokalizacja: dolne środkowe osady formacji Hell Creek, hrabstwo Garfield, Montana (Scannella i Fowler, 2014);
- MOR 3015 - kość łuskowa i mózgoczaszka. Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 3029 - kość ciemieniowa (Scannella i Fowler, 2014; Scannella i in., 2015, ryc. 7.12). Lokalizacja: górne górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 3030 - kryza i kość zaoczodołowa młodego osobnika. Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 3041 - kości łuskowe i mózgoczaszka. Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 3043 - kość łuskowa. Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 3046 - kość ciemieniowa i zębowa. Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 3056 - kość łuskowa i róg nadoczny młodego osobnika. Lokalizacja: górne górne osady formacji Hell Creek (Scannella i Fowler, 2014);
- MOR 3064 - mały (porównywalny z MOR 2951; oszacowana na podstawie długości kości jarzmowej długość podstawowa czaszki to ok. 54 cm - Scannella i in., 2014) młody osobnik, m.in. częściowa kość łuskowa, róg nadoczny (Scannella i Fowler, 2014). Lokalizacja: górne środkowe osady formacji Hell Creek - niżej niż okazy T. ?sp. nov., hrabstwo Garfield, Montana (Scannella i Fowler, 2014);
- MOR 6648 - kość ciemieniowa, łuskowa ?i biodrowa młodego osobnika. Lokalizacja: górne górne osady formacji Hell Creek, hrabstwo Garfield, Montana (Scannella i Fowler, 2014);
- MOR 7037 - kość łuskowa. Lokalizacja: środkowe osady formacji Hell Creek, Montana (Scannella i Fowler, 2014).
= 28 okazów.
Okazy o nieznanym materiale:
- LACM 150076 - duży, niekompletny szkielet zaliczony do Triceratops (Longrich, 2011), nie jest jednak pewne, czy zawiera czaszkę - jest zmontowany w szkielet wraz z LACM 151459 ("GET AWAY TRIKE!", online 2014). Lokalizacja: formacja Lance (Glut, 2006 podał „Laramie”), hrabstwo Niobrara, Wyoming (Glut, 2006, s. 7; LACM, online).
- MOR 622 - częściowa czaszka (Scannella i Fowler, 2014). Lokalizacja: górne osady formacji Hell Creek, Montana (Horner i in., 2011; Scannella i Fowler, 2014);
- MOR 1157 - niekompletna czaszka zaliczona do Triceratops (Holliday, 2009). Lokalizacja: dolne osady formacji Hell Creek, Montana (Horner i in., 2011; Scannella i Fowler, 2014)
- MOR 1129 - częściowa czaszka młodego osobnika (Horner i Goodwin, 2006; Scannella i Fowler, 2014). Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 3016 - rogi nadoczne i fragmenty czaszki. Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR 3053 - m.in. róg nadoczny. Lokalizacja: górne osady formacji Hell Creek, Montana (Scannella i Fowler, 2014);
- MOR P01.1. Lokalizacja: dolne osady formacji Hell Creek, Montana (Horner i in., 2011);
- MPM 51016 (nazwa lokalizacji) - czaszka i szkielet zaliczony do Triceratops (Pearson i in., 2002). Lokalizacja: górne osady formacji Hell Creek, Dakota Północna albo Dakota Południowa (Pearson i in., 2002);
- NDGS 92-19.1 (nazwa lokalizacji) - czaszka i szkielet zaliczony do Triceratops (Pearson i in., 2002) lub szkielet pozaczaszkowy Ceratopsidae indet. (Lyson i Longrich, 2011). Lokalizacja: górne osady formacji Hell Creek, Dakota Północna (Pearson i in., 2002; Lyson i Longrich, 2011);
- NDGS 95-117.1 (nazwa lokalizacji) - czaszka zaliczona do Triceratops (Pearson i in., 2002). Lokalizacja: górne osady formacji Hell Creek, Dakota Północna albo Dakota Południowa (Pearson i in., 2002);
- NDGS 96-23.1 (nazwa lokalizacji) - m.in. czaszka zaliczona do Triceratops (Pearson i in., 2002; Lyson i Longrich, 2011). Lokalizacja: dolne osady formacji Hell Creek, hrabstwo Slope, Dakota Północna (Pearson i in., 2002);
- PTRM v97009 (nazwa lokalizacji) - kryza (Pearson i in., 2002) albo kręgi (Lyson i Longrich, 2011). Lokalizacja: górne osady formacji Hell Creek, hrabstwo Slope, Dakota Północna (Pearson i in., 2002);
- ROM 59423 – co najmniej mózgoczaszka i kłykieć potyliczny, zaliczone do Triceratops (Borkovic, 2013; Paulina-Carabajal i in., 2016);
- ROM 59424 – co najmniej mózgoczaszka i kłykieć potyliczny, zaliczone do Triceratops (Borkovic, 2013; Paulina-Carabajal i in., 2016);
- ROM 59425 – co najmniej kłykieć potyliczny zaliczony do Triceratops (Borkovic, 2013);
- ROM 59426 – co najmniej kłykieć potyliczny, o ogromnych rozmiarach (zob. Torosaurus: Największe ceratopsy) zaliczony do Triceratops (Borkovic, 2013);
- ROM 59502 – co najmniej kłykieć potyliczny zaliczony do Triceratops (Borkovic, 2013); według Daley i Budda (2010) to kambryjski bezkręgowiec Caryosyntrips;
- ROM 59502 – co najmniej mózgoczaszka i kłykieć potyliczny, zaliczone do Triceratops (Borkovic, 2013; Paulina-Carabajal i in., 2016);
- ROM 59423 – co najmniej mózgoczaszka i kłykieć potyliczny, zaliczone do Triceratops (Borkovic, 2013; Paulina-Carabajal i in., 2016);
- SMM P2-A-760 – zaliczony przez Forster (1990) i Andersona (1999). Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Anderson, 1999);
- UCMP 128562 - młody osobnik (Horner i Goodwin, 2008); formacja Hell Creek, hrabstwo Garfield, Montana (UCMP, online);
- UCMP 159223 - niekompletna czaszka, w tym episquamosale (Horner i Goodwin, 2008, s. 142);
- USNM 4805 - zaliczany do Triceratops (Hatcher i in., 1907; Ostrom i Wellnhofer, 1986; CoBabe i Fastovsky, 1987; Anderson, 1999), zawiera m.in. żuchwę i róg nosowy przypominający T. horridus (Hatcher i in., 1907, s. 136-138; Scannella i in., 2015, ryc. 7.7A), do którego został zaliczony przez Borkovica (2013). Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Hatcher i in., 1907, s. 135 i 136);
- USNM 5812 - zaliczony do Triceratops (Forster, 1996a; Lyson i Longrich, 2011), zawiera m.in. róg nosowy (Forster, 1996a, s. 247, 248). Ilustracje: USNM, online. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (USNM, online);
- USNM 6062 - fragmenty czaszki, zaliczone do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Denver, Colorado (Lyson i Longrich, 2011);
- ? USNM 24216 (Holliday, 2009). Baza USNM (online) nie wskazuje takiego numeru, więc jest zapewne pomyłką;
- UWGM A-041-7-4/02 - czaszka zaliczona do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Montana (Lyson i Longrich, 2011);
- YPM 1829 - zaliczony do Triceratops (Lull, 1915; 1933; Ostrom i Wellnhofer, 1986, s. 118). Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Lull, 1915);
- YPM 1833 - zaliczony do Triceratops/Nedoceratops (Lull, 1915; 1933; Ostrom i Wellnhofer, 1986, s. 118; Lyson i Longrich, 2011). Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Lull, 1915);
- YPM 1837 - zaliczony do Triceratops (Lull, 1915; 1933; Lyson i Longrich, 2011). Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Lull, 1915);
- YPM 1838 - zaliczony do Triceratops (Lull, 1915; 1933; Lyson i Longrich, 2011). Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Lull, 1915);
- YPM 2093 - według Forster (1990; 1996a, s. 256) to min. mózgoczaszka Triceratops, lecz według bazy danych YPM (online) jest to odcisk stopy Grallator;
- YPM 3220 – zaliczony do Triceratops sp. (Maiorino i in., 2015; YPM, online).
= 33 okazy.
Zaliczone przez Andersona (1999), nie jest pewne, na jakiej podstawie, być może - tak jak okaz przez niego opisany (UM 17062) - z powodu podobieństwa mózgoczaszki i rzadkości Torosaurus:
- MPM 81.426. Lokalizacja: formacja Hell Creek (Anderson, 1999);
- SMM P64.17.3. Lokalizacja: formacja Hell Creek (Anderson, 1999);
- SMM P66.33.1. Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (Anderson, 1999);
- SMM P60.9.1 – zawiera kość przedszczękową, zaliczoną do Triceratops (Forster, 1993). Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (Anderson, 1999);
- SMM P62.15.2. Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (Anderson, 1999);
- SMM P93.16.1. Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (Anderson, 1999);
- SMM P93.17.1. Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (Anderson, 1999);
- SMM P96.11.1. Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (Anderson, 1999);
- SMM P2-A-579. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Anderson, 1999);
- UM 17062 - zaliczony do Triceratops, gdyż budowa mózgoczaszki przypomina zilustrowane okazy a Torosaurus się różni (Anderson, 1999). Lokalizacja: formacja Hell Creek, hrabstwo McCone, Montana (Anderson, 1999).
= 10 okazów.
Materiał zaliczany czasem do Triceratops o niepewnej przynależności:
- AMNH 5030, zaliczony do Triceratops (Anderson, 1999) lub Ceratopsidae indet. (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, płd.-zach. Lisman, Montana (Anderson, 1999);
- UCMP 140224, zawiera kości łuskowe (Farke i in., 2009) albo kość szczękową zidentyfikowaną jako Triceratops (Wolff, 2007) albo krzyżową (UCMP, online). Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (UCMP, online);
- USNM 2147 - zaliczony do Triceratops (Forster, 1990; 1996a, s. 253; Wolff, 2007), zawiera przynajmniej róg nadoczny (Forster, 1996a, s. 253; USNM, online; Farke, online 2012b). Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (USNM, online; Farke, online 2012b);
- USNM 4708 - zaliczany do Triceratops (Lull, 1915, s. 343; 1933, s. 15; Gilmore, 1920, tabl. 9; Ostrom i Wellnhofer, 1986, s. 117; Butler i Barrett, 2008; Wolff, 2007), konkretniej T. horridus' (Farke, online 2012b), zawiera kość ?ciemieniową (USNM, online), nie jest jednak ujęty w Farke i in. (2009) a Farke (2004) podaje, że to "prawdopodobnie Triceratops".. Ilustracje: Gilmore, 1920, tabl. 9 Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Lull, 1933);
- USNM 8059, zaliczany do Triceratops (Lyson i Longrich, 2011; USNM, online) lub ?Triceratops (Farke, online 2012b). Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (USNM, online);
- YPM 1836 - zaliczony do Triceratops (Lull, 1915; Lyson i Longrich, 2011) lub ?Triceratops (Lull, 1915). Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Lull, 1915).
= 6 okazów. = łącznie: 311 okazów.
Materiał wykluczony
Wybrany materiał wykluczony z Triceratops, głównie z powodu braku elementów diagnostycznych:
- AMNH 5039 - przypisany do Triceratops (Maiorino i in., 2017), zawiera żuchwę i kość ramienną (AMNH, online – 5039; Chinnery, 2004; Maidment i in., 2012);
- BHI 6441 - dolna szczęka zaliczona do T. horridus (Maiorino i in., 2017);
- CCM V2015.7.1 – niekompletna kość ciemieniowa o szerokości 25,3 cm i grubości 1,31-1,5 cm (Wilson i Fowler, 2017). Ilustracje: Wilson i Fowler, 2017. Lokalizacja: ?dolne ?środkowe osady formacji Hell Creek (2015-FPG-023; ‘Jack's Blue Juvie’), hrabstwo Garfield, Montana (Wilson i Fowler, 2017);
- CMN 8598 (=GSC 8598 - Forster, 1996a, s. 247) - przednia część kości nosowych i róg nosowy mierzący ok. 30,5 cm (=12 cali), przypisane do T. prorsus przez Sternberga (1949, s. 43). Jeśli interpretacja Sternberga jako rogu jest prawidłowa to jest to T. prorsus, lecz zapewne nie jest, gdyż przez cały ?róg ciągnie się szew, nieobecny na rogach, lecz między kości nosowymi (por. ryc. 3B w Horner i Goodwin, 2008), zatem jest to raczej niediagnostyczna kość nosowa, być może z niewielkim rogiem. Ilustracje: Sternberg, 1949, tabl. VII. Lokalizacja: formacja Frenchman, okolica Morgan Creek, południowe Saskatchewan (Sternberg, 1949);
- CNM 8864 (?=GSC 8864) - róg nosowy, przypisany do T. prorsus przez Sternberga (1949, s. 43). Jak wyżej, interpretacja Sternberga jako rogu jest zapewne nieprawidłowa, gdyż jest widoczny szew. Lokalizacja: formacja Frenchman, Saskatchewan (Sternberg, 1949);
- FPDM-V-9775 - puszka mózgowa (Sakagami i Kawabe, 2020). Ilustracje: Sakagami i Kawabe, 2020. Lokalizacja: Hell Creek, Marmarth, Dakota Północna (Sakagami i Kawabe, 2020);
- okaz z MGM (obecnie w NBC; wcześniej YPM 1832 – czaszka 21 – por. "4Visualization", online 2016; Bastiaans i in., 2015 oraz Lull, 1915, s. 343) - znacznie zrekonstruowana czaszka, która w czasie transportu rozpadła się na ponad 600 elementów (Schuyf, 1969; Bastiaans i in., 2015; "4Visualization", online 2016). Abstrakt Bastiaansa i in. (2015) różni się od posteru – posiłkując się zdjęciami, tekst tego drugiego pod względem zachowanego materiału wydaje się prawidłowy, zaś rekonstrukcja Forda (online – Triceratops) jest nieprawidłowa. Okaz został zaliczony do Triceratops (Lull, 1915; 1933; Schuyf, 1969; Ostrom i Wellnhofer, 1986, s. 118; Lyson i Longrich, 2011), w tym jako "plezjotyp T. ?brevicornus" a później do T. prorsus, o czym świadczyć mają zamknięte ciemiączka czołowo-ciemieniowe (Bastiaans i in., 2015), lecz są one obecne również u Torosaurus (Farke, 2006) a cienkie fragmenty kryzy (Bastiaans i in., 2015) również wskazują na możliwą przynależność do tego drugiego. Ilustracje: *; "4Visualization", online 2016; Bastiaans, online 2014; Bastiaans i in., 2015; NBC, online; Schuyf, 1969. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Lull, 1915; Bastiaans i in., 2015; Schuyf, 1969);
- MOR 6653 - zawiera dużą kość przedszczękową (Scannella i in., 2014; Scannella, 2020). Może być diagnostyczny. Lokalizacja: dolne środkowe osady formacji Hell Creek, Montana (Scannella i in., 2014);
- MRF v03TRI - szkielet pozaczaszkowy, zaliczony do Triceratops (Lyson i Longrich, 2011). Lokalizacja: formacja Hell Creek, Dakota Północna (Lyson i Longrich, 2011);
- NDGS 1715 – niediagnostyczne elementy czaszkowe i pozaczaszkowe, w tym patologiczne kręgi ogonowe (Canoy Illies i Fowler, 2020);
- PTRM v98017 - kręgi, zaliczone do Triceratops (Lyson i Longrich, 2011), według Pearsona i in. (2002) jest to bliżej nieokreślony ceratops. Lokalizacja: górne osady formacji Hell Creek, hrabstwo Slope, Dakota Północna (Pearson i in., 2002);
- UCM 38473 - róg nosowy, nadoczny i elementy pozaczaszkowe, zaliczone do T. horridus (Carpenter, 1979; Carpenter i Young, 2002) lub Triceratops sp. (Lyson i Longrich, 2011). Lokalizacja: formacja Denver/Laramie, hrabstwo Weld, Colorado (Carpenter, 1979; Carpenter i Young, 2002; Lyson i Longrich, 2011);
- UCMP 138597 - Triceratops według Wolffa (2007), według bazy danych UCMP (online) jest to żuchwa. Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (UCMP, online);
- UCMP 144297 – kości nosowe, jarzmowa (Farke i in., 2009), rostralna i przedszczękowa(?) (Horner i Goodwin, 2008); UCMP online podaje tylko k. szczękową), Triceratops według Wolffa (2007) oraz Goodwina i Hornera (2001). Goodwin i Horner (2001) podają, że róg nosowy jest zakrzywiony, ostry i nieznacznie ściśnięty. Lokalizacja: formacja Hell Creek, hrabstwo Garfield, Montana (UCMP, online);
- USNM 101 - przypisany do Triceratops min. róg nosowy i kość nosowa (Forster, 1996a, s. 248), według bazy danych USNM nie ma takiego okazu;
- USNM 398270 - co najmniej róg nadoczny, przypisany do Triceratops (Borkovic, 2013); według USNM (online) nie ma takiego okazu;
- USNM 4286c - co najmniej kłykieć potyliczny, przypisany do Triceratops (Borkovic, 2013); według USNM (online) nie ma takiego okazu;
- USNM 508495 - fragment czaszki i dolna szczęka zaliczone do Triceratops (Maiorino i in., 2017). Ilustracje: USNM, online. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (USNM, online);
- USNM 7085 - róg nadoczny (USNM, online), przypisany do Triceratops (Borkovic, 2013). Ilustracje: USNM, online.
- USNM 7086 - róg nadoczny (USNM, online), przypisany do Triceratops (Borkovic, 2013). Ilustracje: USNM, online;
- USNM 8027 - róg nadoczny (USNM, online), przypisany do Triceratops (Borkovic, 2013). Ilustracje: USNM, online;
- USNM 8054 - róg nadoczny (USNM, online), przypisany do Triceratops (Borkovic, 2013). Ilustracje: USNM, online;
- ? WCCH, brak numeru (wcześniej UCM 41777; "Pops") – czaszka, żuchwa, 2 kręgi grzebietowe, fragmenty żeber, 10 kręgów ogonowych, kości kulszowe I łonowa, przypisane do T. horridus przez Carpentera i Younga (2002). Kryza jest niekompletna, więc nie można mieć pewności, czy to Triceratops. Jeżeli duży fragment kryzy został umieszczony we właściwym miejscu – tj. jest kością łuskową zawierającą krawędź łączenia z kością ciemieniową, to okaz nie ma charakterystycznego dla Torosaurus latus podłużnego zgrubienia na kości łuskowej. Okaz pochodzi z osadów starszych niż formacja Hell Creek a nawet niż Eotriceratops – tj. sprzed ok. 69-68,5 Ma (Fowler, online 2016; 2017; online 2017) lub 69,6-68,9 Ma (Eberth i Kamo, 2020) (magnetochron 31n). Sugeruje to, że może to nie być Triceratops, lecz inny takson (Fowler oznaczył ten okaz jako „Laramie 'Triceratops'”). Czas i miejsce występowania jest bardziej zbliżone do Ojoceratops fowleri i ?Torosaurus utahensis z Nowego Meksyku (zasięgi czasowe pokrywają się [Fowler, online 2016; 2017]) niż do Eotriceratops z Alberty (nieznacznie późniejszego). Rogi nadoczne są znacznie ciensze niż u ?Torosaurus utahensis, zatem jest to prawdopodobnie Ojoceratops fowleri, którego można traktować jako wczesny gatunek Triceratops. Ilustracje: *; Carpenter i Young, 2002, ryc. 10. Lokalizacja: formacja Laramie, Briggsdale, Colorado (Carpenter i Young, 2002; Lyson i Longrich, 2011; Fowler, online 2016; 2017)
- WMUC 544 – przypisany do T. prorsus przez Hatchera i in. (1907), jako róg nosowy młodego okazu. Jest wydłużony, lecz co nietypowe jego szerokość jest większa niż wysokość. Mógł należeć do młodego Pachycephalosaurus (sensu Horner i Goodwin, 2009) – por. Bakker i in. (2006), Galton i Sues (1983), Goodwin i in. (1998), Sues i Galton (1987). Ilustracje: Hatcher i in., 1907, ryc. 29.
Wątpliwe gatunki
T. albertensis

Nazwany: Sternberg, 1949
Holotyp: CNM 8862 (=GSC 8862) – gigantyczna, prawdopodobnie największa wśród ceratopsów (zob. Torosaurus: Największe ceratopsy), lecz niekompletna czaszka, dwa kręgi grzbietowe i żebra (Sternberg, 1949). Szacowana długość czaszki to minimum ok. 244 cm (=8 stóp). Zachowana część ma 193 cm (Sternberg, 1949). Zależnie od rekonstrukcji przedniej części pyska, długość czaszki mogła wynosić ok. 2,3-2,5 m, a nawet więcej. Okaz opisany w: Sternberg, 1949. Ilustracje: Sternberg, 1949, tabl. VI; Glut, 1997, s. 923; Ford, online - Nedoceratops. Lokalizacja: formacja Scollard, płd. Alberta (Forster, 1996b).
Status: nomen dubium, prawdopodobnie Triceratops
Komentarz: według Forster (1996b) T. albertensis to nomen dubium, lecz na pewno Triceratops, zaś Glut (1997) i Eberth i in. (2013) zaliczyli go do T. horridus. Ostrom i Wellnhofer (1986, s. 144) uznali, że to prawdopodobnie przedstawiciel monotypowego Triceratops, lecz sugerowali, że nie ma pewności do przynależności do rodzaju. „Zaokrąglona bocznie” kość łuskowa, mająca „kształt i proporcje pośrednie między T. eurycephalus a T. flabellatus” (Sternberg, 1949, s. 40), której budowa „leży w zakresie Triceratops” (Forster, 1996b, s. 266) mogą wskazywać, że CNM 8862 to Triceratops, lecz materiał jest słabo zachowany i zdeformowany (Forster, 1996b, s. 266) oraz słabo zilustrowany i opisany, co powoduje, że nie ma pewności co do rzeczywistej morfologii (por. Scannella i Horner, 2010, ryc. 3). Szczególnie tylna część kości łuskowej wydaje się niekompletna, choć kompletność sugeruje podana przez Sternberga liczba episquamosales – 7 (Ford [online - Nedoceratops] zilustrował 3). Nierozwinięte wcięcie jarzmowe sugeruje Triceratops, lecz nie wyklucza ona Torosaurus (zob. Torosaurus: wcięcie jarzmowe) a budowa CNM 8862 nie jest na tyle pewna, aby uznać tą cechę za przesądzającą o identyfikacji. Duża liczba episquamosales (7) jest bardziej typowa dla Torosaurus niż Triceratops (jedyny okaz Triceratops z tak dużą liczbą to MOR 1120 [Scannella i Horner, 2010]). Kość łuskowa została określona jako „gruba” (Sternberg, 1949, s. 38), więc prawdopodobnie jest to rzeczywiście Triceratops. Niekompletne, długie na ponad 72 cm rogi nadoczne (od oczodołu - Sternberg, 1949, s. 40) sugerować mogą, że to nie T. prorsus, lecz z uwagi na wielkie rozmiary okazu, powodujące, że stosunek długości rogu do długości czaszki jest dość mały, nie można wykluczyć przynależności do tego gatunku. Ford (online - Nedoceratops) przypisuje CNM 8862 do Nedoceratops (jako N. albertensis), być może z powodu prostych, skierowanych ku tyłowi rogów nadocznych. Budowa ta zdaniem Forster (1996b) wynika zapewne z deformacji pośmiertnej a według Ostroma i Wellnhofera (1986, s. 143) częściowo jest wynikiem nieprawidłowego ułożenia czaszki, której przód zapewne powinien zostać ukierunkowany 15-20 stopni w dół. Inaczej twierdził Sternberg (1949, s. 40), według którego kąt zwrócenia rogów jest naturalny, bo czaszka tkwiła w skale na boku.
T. alticornis
Nazwany: (Marsh, 1887) Hatcher 1907 vide Hatcher, Marsh i Lull, 1907
Holotyp: USNM 4739 - dwa połączone rogi nadoczne. Często podaje się nr 1871 lub 1871E (Hatcher i in., 1907; Ostrom i Wellnhofer, 1986); Lull (1933, s. 16) podaje, że jest to nr innego okazu z tej lokalizacji. Ilustracje: * = Hatcher i in., 1907, ryc. 106; Glut, 1997, s. 919; USNM, online. Lokalizacja: formacja Denver, miasto Denver, Lakewood Gulch, Colorado (Ostrom i Wellnhofer, 1986; Carpenter, 2006).
Status: nomen dubium, niekoniecznie Triceratops (Forster, 1996b).
Komentarz: słynny okaz, nazwany Bison alticornis przez Marsha (1887), który sądził początkowo, że skamieniałość pochodzi z późniejszych warstw.
T. eurycephalus
Nazwany: Schlaikjer, 1935
Holotyp: MCZ 1102 - dość kompletna czaszka o długości ok. 138,6 cm (Schlaikjer, 1935). Ilustracje: *; Schlaikjer, 1935; Glut, 1997, s. 925; Hudson, online 2008; "Oregon Hiker", online 2007b; Estes, online 2013. Lokalizacja: formacja Lance, hrabstwo Goshen, Wyoming (Schlaikjer, 1935).
Status: nomen dubium - młodszy synonim T. horridus lub T. ?sp. nov. albo ważny gatunek Triceratops
Komentarz: według Forster (1996b) mały kłykieć potyliczny i delikatne kości żuchwy nie przypominają Triceratops a fragmentaryczna kryza z brakiem różków wykluczają potwierdzenie, że jest to ten rodzaj. Kryza jest, zgodnie z nazwą (eurycephalus to „szerokoczaszkowy“) szczególnie szeroka – ma 129,7 cm (Schlaikjer, 1935). Kość udowa jest długa, lecz smukła. Tylny koniec kości łuskowej ma nietypowy, zaokrąglony kształt (Schlaikjer, 1935). Mogą to być cechy diagnostyczne.
T. galeus

Nazwany: Marsh, 1889b
Holotyp: USNM 2410 - róg nosowy (Forster, 1996b) i "inne pozostałości, prawdopodobnie z tego samego przedziału (horizon)" (Marsh, 1889b), opisane w: Hatcher i in., 1907, s. 132. Ilustracje: *; Hatcher i in., 1907, ryc. 111. Lokalizacja: formacja Denver lub Arapahoe, Henderson, Colorado (Ostrom i Wellnhofer, 1986; Carpenter i Young, 2002).
Status: nomen dubium, niekoniecznie Triceratops (Forster, 1996b).
Komentarz: okaz bywa zaliczany do Torosaurus (Forster, 1990, s. 123; Ford, online - Torosaurus), lecz wątpliwe, by był na tyle diagnostyczny (por. Torosaurus: Łukowaty ślad naczynia na rogu nosowym).
T. maximus
Nazwany: Brown, 1933
Holotyp: AMNH 5040 - syncervical, 8 kręgów (5 szyjnych i 3 grzbietowe) i żebra szyjne (Brown, 1933). Ilustracje: Brown, 1933; Glut, 1997, s. 927. Lokalizacja: dolne dolne osady formacji Hell Creek (por. Brown, 1907, s. 829; 1933; Scannella i in., 2014), hrabstwo Garfield, Montana (Brown, 1933).
Status: nomen dubium, niekoniecznie Triceratops (Forster, 1996b).
Komentarz: jak wskazuje nazwa, jest to olbrzymi okaz. Zob. Torosaurus: Największe ceratopsy.
T. flabellatus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
.
Holotyp to YPM 1821 to czaszka i szkielet pozaczaszkowy opisane w: Hatcher i in., 1907, s. 15 i n. oraz 143 i n. Ilustracje: Hatcher i in., 1907, tabl. XLIV-XLVI; Lull, 1934, ryc. 2 i 3; Glut, 1997, s. 924; "Oregon Hiker", online 2007a. Lokalizacja: formacja Lance, hrabstwo Niobrara, Wyoming (Hatcher i in. 1907; Ostrom i Wellnhofer, 1986);
YPM 1821 to na pewno Triceratops, gdyż mają praktycznie kompletną kryzę z diagnostyczną morfologią. Jest to spory, ale dość młody osobnik (np. Hatcher i in., 1907; Longrich i Field, 2012). Otwarte ciemiączko czołowo-ciemieniowe (Hatcher i in., 1907, s. 145; Lull, 1934, s. 442, Forster, 1996b, s. 265; contra Lehman, 1998, s. 903) wskazywałyby na T. horridus (albo okazy „pośrednie” - tj. T. ?flabellatus), lecz wynikać to może z niedojrzałości okazu. Czaszka w okolicy rogu nosowego jest niekompletna (Hatcher i in., 1907). Utrudnia to przyporządkowanie do któreś z form triceratopsa. Stratygraficznie YPM 1821 znajduje się wyżej od okazów typowych T. prorsus i T. horridus (Lull, 1915; Ostrom i Wellnhofer, 1986), co wskazywałoby, że to T. prorsus, jednak podobną pozycję ma wiele okazów uznawanych za T. horridus (co nasuwa podejrzenie, że pozycja stratygraficzna holotypu T. prorsus została nieprawidłowo określona albo hipoteza o rozdzieleniu czasowym gatunków jest błędna). Róg nosowy YPM 1821 mógł wykazywać morfologię T. horridus (Lull, 1934, ryc. 2), lecz Lull (1933, s. 121) stwierdza też, że równie dobrze mógł mieć róg taki jak u typowego okazu T. brevicornus (=T. prorsus). YPM 1821 mógł mieć, jak T. horridus i forma „pośrednia”, długie rogi nadoczne. Marsh (1889b) stwierdził, że rogi mają około trzy stopy (ok. 91 cm), co zostało zapewne powtórzone przez Dodsona (1996, s. 63), podającego 90 cm. 71 cm podał Borkovic (2013), przy czym autor ten mierzył zapewne zrekonstruowane rogi (Lull, 1934) i stosował punkty pomiarów odmienne od większości badaczy, skutkujące tym, że podane przez niego wartości są o kilka cm mniejsze. Rogi zostały zilustrowane (np. Marsh, 1889c), choć ilustracje z publikacji Marsha i Hatchera ukazują często uzupełnioną morfologię (np. USNM 4928). Hatcher i in. (1907, s. 145) podali jednak, że odnaleziony został tylko lewy róg i to w złym stanie. Według Lulla (1934) róg nie został przez niego zlokalizowany i mógł ulec zniszczeniu a obecne w okazie po jego złożeniu niedługo przed pisaniem pracy są zrekonstruowane. Lull (1933, s. 121) opisał jednak podstawy rogów. Także Forster (1996b, s. 265) podała, że rogi nie są zachowane.
YPM 1821 nie został wystarczająco dobrze zilustrowany, morfologia wymaga jeszcze dokładniejszego porównania z innymi okazami.
T. serratus
Nazwany: Marsh, 1890a
Holotyp: YPM 1823 – czaszka należąca do dość młodego okazu (np. Hatcher i in., 1907; Longrich i Field, 2012), opisana w: Hatcher i in., 1907, s. 122 i n. Ilustracje: Hatcher i in., 1907, tabl. XXVII-XXIX; Glut, 1997, s. 920 i 923. Lokalizacja: formacja Lance, Lightning Creek, hrabstwo Niobrara, Wyoming (Ostrom i Wellnhofer, 1986).
Status: nomen dubium, na pewno Triceratops.
Komentarz: jest to z pewnością Triceratops, gdyż wykazuje diagnostyczną dla niego budowę kryzy (Hatcher i in., 1907). Forster (1996b) uznała go za młodszy synonim T. horridus, gdyż ma według niej długie rogi nadoczne, otwarte ciemiączka i „S-kształtny” pysk; za nią Anderson (1999). Longrich i Field (2012, ryc. 5) oraz Longrich (2014) zaliczyli jego holotyp do T. prorsus, zaś Maiorino i in. (2013b) w suplemencie do T. horridus, ale w rycinach widnieje jako T. prorsus. Zatem T. serratus to potencjalny młodszy synonim T. horridus albo starszy lub młodszy synonim T. prorsus. T. serratus i T. prorsus zostały ustanowione w tej samej publikacji (Marsh, 1890a), więc rozstrzygnięcie o tym, który powinien być uznany za starszy synonim należy do pierwszego autora, który w pracy opublikowanej zgodnie z wymaganiami International Code of Zoological Nomenclature (ICZN) zacytuje obie nazwy i wybierze jedną z nich (ang. First Reviser) (art. 24.2.1. ICZN) i wydaje się, że powinien zostać wybrany T. prorsus, gdyż był częściej uznawany za ważny (Recommendation 24A ICZN). Publikacja Longricha i Fielda (2012) nie spełnia warunków art. 24.2.1. ICZN. Otwarte ciemiączko czołowo-ciemieniowe (Hatcher i in., 1907, s. 125; contra Lehman, 1998, s. 903) wskazywałyby na T. horridus albo T. ?sp. nov., lecz wynikać może z niedojrzałości okazu. Czaszka w okolicy rogu nosowego jest niekompletna (Hatcher i in., 1907), co utrudnia to przyporządkowanie ich do któreś z form triceratopsa. Róg nosowy nie zachował się, lecz według Lulla (1933, s. 120) – co budzi jednak poważne wątpliwości – nie mógł być masywny ani długi, co sugeruje T. horridus. Dolna krawędź górnej części kości nosowej jest prosta, co sugeruje T. prorsus (Scannella i in., 2014), lecz kość jest pęknięta i może być zniekształcona – zbyt wywinięta ku górze (por. ilustracje w Glut, 1997, s. 923; Hatcher i in., 1907, tabl. XXVII). Choć rogi nadoczne są zachowane w około (Hatcher i in., 1907, s. 125) lub mniej niż połowie (Lull, 1933, s. 120), Forster (1996b) zakodowała je jako długie a Borkovic (2013) podał ich długość, która mieści się w zakresie wspólnym dla obu gatunków. Analiza kości łuskowych Maiorino i in. (2013b) wykazała, że okaz ten mieści się w zakresie morfologicznym T. prorsus, choć do T. horridus nie jest mu daleko. Okaz nie został wystarczająco zilustrowany i wymaga dokładniejszego zbadania. W świetle opublikowanych danych należy więc uznać T. serratus za nomen dubium.
T. sulcatus
Nazwany: Marsh, 1890b
Holotyp: USNM 4276 - rogi nadoczne (Forster, 1996b), Marsh (1890b) pisze o kompletnej czaszce (w Hatcher i in. [1907] mowa jest o fragmentarycznej czaszce, tak samo Lull [1933, s. 128]), kręgach i innych skamieniałościach. Maidment i in. (2012) podają wymiary kości ramiennej. Maiorino i in. (2017) podają numer 4726 jako dolną szczękę, lecz pod tym numerem widnieje w katalogu USNM (online) Apatosaurus. Okaz opisany w: Hatcher i in., 1907, s. 133-134. Ilustracje: Hatcher i in., 1907, ryc. 112; Ford, online - Triceratops; USNM, online. Lokalizacja: formacja Lance, Lance Creek, hrabstwo Niobrara, Wyoming (Ostrom i Wellnhofer, 1986).
Status: nomen dubium, niekoniecznie Triceratops (Forster, 1996b).
Komentarz: okaz ma głęboką bruzdę na rogu nadocznym (sulcus), od której wziął swoją nazwę. Bruzda znajdowała się na tylnej części rogu, który zaginął. Kilka innych okazów ma bardziej lub mniej rozwiniętą bruzdę na przedniej części rogów, lecz nie uważa się tej cechy za diagnostyczną (Hatcher i in., 1907, s. 133-134; Lull, 1933, s. 129). Dwie bruzdy ma Torosaurus latus MOR 1122 (Farke, 2006).
Media


{kind=link}
{kind=link}
{kind=link}

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Triceratops jest jednym z najbardziej znanych dinozaurów. Można ustawić go w jednym rzędzie z takimi dinozaurzymi sławami, jak Tyrannosaurus, Stegosaurus i Diplodocus. Jest bohaterem licznych filmów fabularnych, popularnonaukowych czy rysunkowych, a także książek i atlasów o tematyce mezozoicznej. Bardzo często przedstawia się go jako głównego oponenta tyranozaura (dawniej także m.in. allozaura, który żył wiele milionów lat wcześniej) i jednocześnie jako jedynego dinozaura, który jest w stanie się przed nim obronić (czasami za takiego uważa się również Ankylosaurus).
Można powiedzieć, że w kulturze masowej jest reprezentantem ceratopsów, a także jednym z symboli ery dinozaurów w ogóle. Jest przez to jednym z najszerzej rozpoznawalnych na świecie dinozaurów. Często ludzie widząc wizerunek trójrogiego ceratopsyda myślą, że to właśnie Triceratops, wbrew intencjom autora rekonstrukcji. Gatunek T. prorsus był również przez wiele lat symbolem witryny D.com! od momentu jej założenia w 2005 roku przez Łukasza Czepińskiego – rekonstrukcja jego czaszki autorstwa Tomasza Rutkowskiego widniała m.in. na stronie głównej. Także obecnie logiem D.com! jest Triceratops, autorstwa Roberta Bronowicza. Triceratopsa nie może zabraknąć w żadnym szanującym się dinoparku czy pozycji książkowej dla najmłodszych (np. kultowe w Polsce "Dinozaury" DeAgostini z lat 1994-1996), a jeśli chodzi o kino i telewizję, triceratops jest jednym z najczęściej "angażowanych" dinozaurów.
Kino
Jednym z pierwszych filmów pełnometrażowych, a może nawet w ogóle pierwszym, w którym można obejrzeć Triceratops jest pierwotna ekranizacja książki Arthura Conan Doyle’a pt. "Zaginiony Świat"[3] (The Lost World) pod tym samym tytułem z 1925 roku. Film ten jest jednym z prekursorów efektów specjalnych – animatroniki. W jednej ze scen widać pasącego się triceratopsa z młodym. Ich spokój przerywa niespodziewane pojawienie się dużego teropoda, prawdopodobnie Allosaurus[4]. Tym razem do walki nie dochodzi, mięsożerca widocznie nie chce ryzykować i odchodzi, przegoniony przez samicę triceratopsa[5]. Co ciekawe, w innej scenie dochodzi do walki dużego teropoda (prawdopodobnie Allosaurus lub Tyrannosaurus) z ceratopsydem wyglądającym identycznie jak Agathaumas z ryciny Charlesa R. Knighta. Tym razem drapieżnik nie daje żadnych szans roślinożercy[6], a po jego upolowania w jego szczęki wpada także nieostrożny Pteranodon.
W 1966 roku miał premierę głośny film przygodowy pt. "Milion Lat Przed Naszą Erą"[7] (One Million Years B.C.) z piękną Raquel Welch w roli głównej. Akcja toczyła się – zgodnie z tytułem – ponad milion lat temu i opowiadała o losach ponętnej dzikuski. Co ciekawe, ówcześni ludzie jaskiniowi (bez większej charakteryzacji, wyglądający jak ludzie współcześni) żyli obok… dinozaurów, będąc co chwila przez nich nękani. W jednej ze scen[8] nasza blondwłosa piękność, spacerująca z jednym z plemiennych myśliwych, napotyka na żerującego triceratopsa. Uciekając przed nim, wpadają na Ceratosaurus, który toczy z triceratopsem zażarty pojedynek. Teropod przegrywa go, gdy triceratops śmiertelnie przebija jego podbrzusze swymi rogami.
Kolejnym filmem o podobnej konwencji, jest słabo znany w Polsce "Ostatni Dinozaur"[9] (The Last Dinosaur) japońsko-amerykańskiej produkcji z 1977 roku. Tym razem akcja rozgrywa się w czasach współczesnych – a być może zważywszy na ukazaną w nim zaawansowaną technologię – w niedalekiej przyszłości. Tym razem ekspedycja naukowa (z piękną dziennikarką na czele) wyrusza na Antarktydę, gdzie odkrywa nowy ląd, ukryty pod lodowcem. Okazuje się, że żyją tam ludzie pierwotni wraz z dinozaurami. W jednej ze scen[10] dochodzi do walki Triceratops z Tyrannosaurus. Mimo początkowej przewagi triceratopsa, który przebija tyranozaura swoimi rogami, a nawet kąsa go swym dziobatym pyskiem, traci inicjatywę na rzecz T. rex, który przewraca go i zagryza. Modele dinozaurów wykonane są w stylu japońskiej Gojiry (Godzilli).
Wreszcie nastał 1993 rok i na ekrany kin całego świata wszedł słynny "Park Jurajski" Stevena Spielberga. Obraz ten zmienił diametralnie postrzeganie dinozaurów przez laików oraz wywołał istny boom na dinozaury wśród dzieci i młodzieży. Triceratops wystąpił we wszystkich częściach trylogii "Park Jurajski". Najbardziej znana scena z triceratopsem miała miejsce w 1. części megaprzeboju z 1993 roku, kiedy to naukowcy dr Alan Grant i dr Ellie Sattler, oprowadzani po tytułowym parku, natykają się na chorą samice triceratopsa. Była ona grana przez animatronicznego robota[11] w skali 1:1, stworzonego przez specjalistów z ILM (Industrial Light & Magic George’a Lucasa). Triceratops był także kilkukrotnie pokazywany w trylogii stadnie, przy wodopoju – tym razem jego modele były stworzone komputerowo.
Telewizja
Do walki Triceratops z Tyrannosaurus[12] dochodzi także w popularnonaukowym serialu dokumentalnym pt. "Dinozaury" (The Dinosaurs) z 1992 roku. I tym razem przegrywa triceratops, zgubiony przez brawurę. Jego nieudana próba przegonienia samicy tyranozaura z młodymi kończy się dla niego fatalnie, a dla matki małych tyranozaurów jedynie stratą jednego zęba i niegroźną szramą na brzuchu. Rysunkowe i bardzo dokładne animacje prezentowane w tym serialu dokumentalnym uchodzą dziś za kultowe.
Co ciekawe, bodaj najgłośniejszy serial paradokumentalny o dinozaurach – "Wędrówki z dinozaurami" (Walking with Dinosaurs) – produkcji BBC z 1999 roku, tylko na chwilę pokazuje triceratopsa, w dodatku martwego. Ma to miejsce w ostatnim odcinku oryginalnej serii pt. "Śmierć dynastii" (jednym z bohaterów odcinka jest Torosaurus, który może być synonimem Triceratops). Nieco więcej uwagi triceratopsowi poświęcają autorzy konkurencji – When Dinosaurs Roamed America ("Wędrówki Dinozaurów" z 2001 roku), gdzie ukazane jest w drugim odcinku pasące się stado tego dinozaura rogatego.
Dinozaury cieszyły się uznaniem wśród najmłodszych od dawna. Bardzo znanym, także w Polsce, jest serial fabularny pt. "Zaginiony Ląd – Dolina Jurajska"[13] (Land of the Lost) z lat 1991-1992. Opowiada o rodzinie Parkerów, którzy wyruszając na wakacje wpadli w wir czasoprzestrzenny, który przeniósł ich do prehistorycznego świata[14]. Jednym z najczęściej występujących dinozaurów w serialu jest właśnie Triceratops, co ciekawe z ułamanym jednym rogiem nadocznym.
Z seriali i filmów animowanych warto wspomnieć o takich kreskówkach, jak seria "Pradawny Ląd"[15] (znany również jako "Zanim Ruszył Czas"; ang. The Land Before Time) , "Dino-Riders"[16][17] (1988), "Dink, Mały Dinozaur"[18][19] (Dink, the Little Dinosaur) z lat 1989-1991, "Mała Syrenka"[20] (odcinek 27. pt. "Kraina Dinozaurów") z lat 1992-1994, "Niebezpieczne Dinozaury"[21][22] (Extreme Dinosaurs) z lat 1997-1998, w których to triceratops odgrywał mniejszą lub większą rolę.
Muzyka
Odlew czaszki BHI 4772, zrekonstruowanej jako Triceratops, ukazał się na okładce jednego z albumów znanego zespołu grungowego Alice in Chains (The Devil Put Dinosaurs Here). Ukryto tam podobno drugą czaszkę, która w połączeniu z pierwszą tworzy wizerunek przypominający diabła (tytuł zaś odnosić się ma do forsowanej przez niektórych idei, jakoby kości dinozaurów zostały zamieszczone w ziemi przez Szatana, aby odwieść ludzi od wiary)[23]. Triceratops to również nazwa japońskiego zespołu rockowego (znanego również jako Tricera[24]), założonego w 1996 roku.
Zabawki
Triceratops jest naturalnie jednym z najczęściej odwzorowywanych modeli dinozaurów dla najmłodszych. Niektóre gumowe figurki tylko symbolicznie odwzorowują triceratopsa (jak zresztą pozostałe dinozaury), lecz można spotkać modele lepszej jakości, także służące bardziej jako modele redukcyjne, aniżeli zabawki dla dzieci. Również fani klocków Lego mogą się czuć usatysfakcjonowani, bowiem duński potentat światowego przemysłu zabawkowego wydał kilka zestawów[25][26][27][28][29] z triceratopsem.
Ocena rekonstrukcji
Wydaje się, że (niezamierzenie) najczęściej ilustrowanym gatunkiem jest T. ?sp. nov. (długie rogi nadoczne i długi róg nosowy), choć jest najrzadszy w zapisie kopalnym, drugi w kolejności jest T. horridus (długie rogi nadoczne i krótki róg nosowy) a T. prorsus (krótkie rogi nadoczne i długi róg nosowy) jest raczej rzadko przedstawiany. Wynika to zapewne z przyzwyczajenia do długorogich form i mniejszej atrakcyjności wizualnej Triceratops z krótkimi rogami nadocznymi (postrzeganego zapewne jako mniej groźnego – słabiej uzbrojonego). Podobnie nieświadome jest też z pewnością częste ilustrowanie jako dojrzałych niezupełnie dorosłych okazów – z dużymi, trójkątnymi różkami na brzegu kryzy i zakrzywionymi ku górze końcami rogów nadocznych. Notorycznym błędem, także w rekonstrukcjach powstałych po 2009 roku, są ustawione pionowo i opierające się na pięciu palcach, często „słoniowate” przednie kończyny triceratopsa. Ponadto ilustracje często nie uwzględniają znanych odcisków skóry i zwłaszcza pokrycia większości czaszki keratyną – niemal wszystkie ilustracje przedstawiają pokryty takim samymi jak reszta ciała łuskami łeb triceratopsa. Jedne z najlepszych rekonstrukcji wykonano na potrzeby gry Saurian[30][31].
Spis gatunków
Taksonomia za źródłami cytowanymi w samym spisie gatunków lub uzasadnieniami i cytowaniami powyżej. Kombinacje T. mortuarius i T. silvestris za Olshevskym (online 2001). Torosaurus latus uznano za odrębny rodzaj i gatunek – zob. opis.
| Triceratops | Marsh, 1889b | |
|---|---|---|
| = Sterrholophus | Marsh, 1891a | |
| = Nedoceratops | Ukrainsky, 2007 | |
| = Diceratus | Mateus, 2008 | |
| = "Diceratops" | Lull vide Hatcher, 1905 [zajęta Förster, 1869] | |
| = Ugrosaurus | CoBabe i Fastovsky, 1987 | |
| ??= Ojoceratops | Sullivan i Lucas, 2010 | |
| ??= Tatankaceratops | Ott i Larson, 2010 | |
| T. horridus | (Marsh, 1889a) Marsh, 1889b | |
| = Ceratops horridus | Marsh, 1889a | |
| = T. calicornis | Marsh, 1898 | |
| = T. elatus | Marsh, 1891b | |
| = T. obtusus | Marsh, 1898 | |
| = T. "ingens" | Marsh vide Lull, 1915 | nomen nudum |
| = Nedoceratops hatcheri | (Lull vide Hatcher, 1905) Ukrainsky, 2007 | |
| = Diceratus hatcheri | (Lull vide Hatcher, 1905) Mateus, 2008 | |
| = "Diceratops" hatcheri | Lull vide Hatcher, 1905 | |
| = Triceratops hatcheri | (Lull vide Hatcher, 1905) Lull, 1907 | |
| = Triceratops ("Diceratops") hatcheri | (Lull vide Hatcher, 1905) Lull, 1933 | |
| = Ugrosaurus olsoni | CoBabe i Fastovsky, 1987 | |
| ??= Ojoceratops fowleri | Sullivan i Lucas, 2010 | |
| T. prorsus | Marsh, 1890a | |
| = Agathaumas prorsus | (Marsh, 1890a) Lydekker, 1893 | |
| = T. brevicornus | Hatcher, 1905 | |
| ??= Tatankaceratops sacrisonorum | Ott i Larson, 2010 | |
| T. flabellatus | Marsh, 1889b | nomen dubium? |
| = Sterrholophus flabellatus | (Marsh, 1889b) Marsh, 1891a | |
| ?T. albertensis | Sternberg, 1949 | nomen dubium |
| ??T. alticornis | (Marsh, 1887) Hatcher, 1907 vide Hatcher, Marsh i Lull, 1907 | nomen dubium |
| = "Ceratops" alticornis | (Marsh, 1887) Marsh, 1889b | |
| = "Bison" alticornis | Marsh, 1887 | |
| = Polyonax alticornis | (Marsh, 1887) Cope, 1889 | |
| ?T. eurycephalus | Schlaikjer, 1935 | nomen dubium? |
| ??T. galeus | Marsh, 1889b | nomen dubium |
| ??T. maximus | Brown, 1933 | nomen dubium |
| T. mortuarius | (Cope, 1874) Kuhn, 1936 | = Polyonax mortuarius |
| T. serratus | Marsh, 1890a | nomen dubium |
| T. silvestris | (Cope, 1872) Kuhn, 1936 | = Agathaumas silvestris |
| ??T. sulcatus | Marsh, 1890a | nomen dubium |
| T. utahensis | (Gilmore, 1946) Longrich, 2014 | = ?Torosaurus utahensis |
Bibliografia
- 225 Magazine (online 2017) https://www.225batonrouge.com/things-to-do/art-exhibits-concerts-baton-rouge-may-2017-2
- "4Visualization" (online 2016) http://4visualization.eu/triceratops-schedel-reconstructie/
- AMNH (online) http://www.amnh.org/dinosaurs
- AMNH (online - 972) http://research.amnh.org/paleontology/search.php?action=detail&specimen_id=52842
- AMNH (online - 5028) http://research.amnh.org/paleontology/search.php?action=detail&specimen_id=47762
- AMNH (online - 5039) http://research.amnh.org/paleontology/search.php?action=detail&specimen_id=47774
- Alexander, R. McN. (1985) "Mechanics of posture and gait of some large dinosaurs" Zoological Journal of the Linnaean Society, 83(1), 1-25. doi:10.1111/j.1096-3642.1985.tb00871.x
- "ALEXCOTTERELL" (online 2014) https://cptsalearningtoteach.wordpress.com/2014/09/30/science-day-thinktank-week-4/
- Anderson, J.F., Hall-Martin, A. & Russell, D.A. (1985) "Long-bone circumference and weight in mammals, birds and dinosaurs" Journal of Zoology, 207(1), 53-61. doi:10.1111/j.1469-7998.1985.tb04915.x
- Anderson, J.S. (1999) "Occipital condyle in the ceratopsian dinosaur Triceratops, with comments on body size variation" Contributions from the Museum of Paleontology, the University of Michigan, 30(8), 215-231.
- anonim (2017) "Mesozoic mail" Prehistoric Times, 121, 6-7.
- "Anselmo" (online 2010) http://www.flickr.com/photos/astrangerinthealps/4864860172
- Anton, J. (2000) "Ceratopsian crests as acoustic devices" Journal of Vertebrate Paleontology, 20(3 - suplement), 27A [abstrakt]
- Arbour, V.M., Burns, M.E., Sissons, R.L. (2009) "A redescription of the ankylosaurid dinosaur Dyoplosaurus acutosquameus Parks, 1924 (Ornithischia: Ankylosauria) and a revision of the genus" Journal of Vertebrate Paleontology, 29(4), 1117-1135. doi:10.1671/039.029.0405
- Arnold, J. (online 2014) http://authorjoelarnold.blogspot.com/2014/06/grand-river-museum-lemmon-sd-2014.html
- ASU News (online 2009) http://www.news.appstate.edu/2009/02/06/geologyf-triceratops-battles/
- ASU, online 2012 https://asunow.asu.edu/node?page=620
- Bailleul, A.M. & Horner, J. R. (2016) "Comparative histology of some craniofacial sutures and skull-base synchondroses in non-avian dinosaurs and their extant phylogenetic bracket" Journal of Anatomy, 229, 252-285. doi:10.1111/joa.12471
- Bakker, R.T., Sullivan, R.M., Porter, V., Larson, P. & Saulsbury, S.J. (2006) "Dracorex hogwartsia, n. gen., n. sp., a spiked, flat-headed pachycephalosaurid dinosaur from the Upper Cretaceous Hell Creek Formation of South Dakota" New Mexico Museum of Natural History and Science Bulletin, 35, 331-345.
- Bamforth, E. L. (2018) "The Triceratops 'Baby Book': An Update on Juvenile Ceratopsians from the Frenchman Formation (66 Ma) of Saskatchewan, Canada" Canadian Palaeontology Conference (Proceedings Volume No. 15), Saskatoon, Saskatchewan [abstrakt zamieszczony na https://www.researchgate.net/publication/327845666_The_Triceratops_'Baby_Book'_An_Update_on_Juvenile_Ceratopsians_from_the_Frenchman_Formation_66_Ma_of_Saskatchewan_Canada ]
- Bamforth, E.L (online 2019a) https://twitter.com/EL_Bamforth/status/1114207351509344257
- Bamforth, E.L (online 2019b) https://twitter.com/EL_Bamforth/status/1116755549658464256
- Barrick, R.E., Stoskopf, M.K. Marcot, J.D. Russell, D.A. & Showers, W.J. (1998) "The thermoregulatory functions of the Triceratops frill and horns: heat flow measured with oxygen isotopes" Journal of Vertebrate Paleontology, 18(4), 746-750. doi:10.1080/02724634.1998.10011103
- Barton, T. (online 2004) http://www.flickr.com/photos/7230309@N05/3639681176
- Bastiaans, D. (online 2014) https://www.facebook.com/media/set/?set=a.966940516655206.1073741841.100000177993122&type=3
- Bastiaans, D., Guliker, M.D. & Schulp, A.S. (2015) "Reconstruction of a historically interesting Triceratops ‘?brevicornus’ from the Lance Formation, Wyoming, USA" 13th Annual Meeting of the European Association of Vertebrate Palaeontologists, Opole, Poland, 8-12 July 2015, 145. https://www.researchgate.net/publication/280101762_Reconstruction_of_a_Historically_Interesting_Triceratops_brevicornus_From_the_Lance_Formation_Wyoming_USA
- Beach, A. (2011a) "Triceratops and Torosaurus synonymy: an evaluation of two BYU specimens" 25th annual Student Research Conference, Brigham Young University [abstrakt zamieszczony na http://cpms.byu.edu/about/spring-research-conference/submitted-abstracts/?yr=2011&id=436 ]
- Beach, A. (2011b) "Triceratops and Torosaurus synonymy: an evaluation of two large specimens from Brigham Young University" Journal of Vertebrate Paleontology 31(5, supl.), 68 [abstrakt]
- Bell, P., Hendrickx, C., Pittman, M., Kaye, T. & Mayr, G. (2022) "The Exquisitely Preserved Dinosaur Skin of Psittacosaurus and the Scaly Skin of Ceratopsian Dinosaurs" PREPRINT (Version 1) Research Square doi:10.21203/rs.3.rs-1374507/v1
- Benson, R.B.J., Campione, N.E., Carrano, M.T., Mannion, P.D., Sullivan, C., Upchurch, P. & Evans, D.C. (2014) "Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage" PLoS Biology, 12(5), e1001853. doi:10.1371/journal.pbio.1001853
- Benson, R.B.J., Hunt, G., Carrano, M.T. & Campione, N. (2017) "Data from: Cope's rule and the adaptive landscape of dinosaur body size evolution" Dryad, Dataset. doi:10.5061/dryad.1t3r4
- BHI (online 2015) https://twitter.com/bhigr_ep/status/573159847065083905
- BHI (online a) http://www.bhigr.com/store/product.php?productid=889
- BHI (online b) http://www.bhigr.com/store/product.php?productid=401
- BHI (online c) http://www.bhigr.com/store/product.php?productid=77
- Biles, J. (online 2015) http://cjonline.com/news-local/2015-09-07/rare-dinosaur-skull-being-prepared-exhibition#
- "Blomman87" (online 2017) https://blomman87.deviantart.com/art/New-species-of-ceratopsian-672990527
- BMRP (online 2010) https://www.facebook.com/photo.php?fbid=458745928704&set=a.392928023704.163564.163053698704&type=1&relevant_count=1
- Boatman, E.M. (2012) "Electron Microscopy and Analytical X-ray Characterization of Compositional and Nanoscale Structural Changes in Fossil Bone" dysertacja doktorska, University of California, 1-164.
- Borkovic, B. (2013) "Investigating Sexual Dimorphism in Ceratopsid Horncores" praca magisterska, University of Calgary, 1-293.
- Britannica (online 2010) http://blogs.britannica.com/2010/09/torosaurus-is-triceratops-5-questions-for-paleontologist-john-scannella [kopia zdjęcia: https://s-media-cache-ak0.pinimg.com/originals/b5/c2/a5/b5c2a52f0f452c8b2652ebc59c4c11ab.jpg ]
- Brown, A.J. & Smith, N. (2017) "Osteohistological evaluation of the squamosal in Triceratops horridus" Society Of Vertebrate Paleontology 77th Annual Meeting, Meeting Program & Abstracts, 88. [abstrakt]
- Brown, B. (1907) "The Hell Creek beds of the Upper Cretaceous of Montana: their relation to contiguous deposits, with faunal and floral lists, and a discussion of their correlation" Bulletin of the American Museum of Natural History, 23, 823-845. http://hdl.handle.net/2246/1936
- Brown, B. (1917) "A complete skeleton of the horned dinosaur Monoclonius, and description of a second skeleton showing skin impressions" Bulletin of the American Museum of Natural History, 37, 281-306. http://hdl.handle.net/2246/1336
- Brown, B. (1933) "A gigantic ceratopsian dinosaur, Triceratops maximus, new species" American Museum novitates, 649: 1-9. http://hdl.handle.net/2246/2064
- Brown, B. & Schlaikjer, E.M. (1940) "The structure and relationships of Protoceratops" Annals of the New York Academy of Sciences, 40, 133-266. doi:10.1111/j.2164-0947.1940.tb00068.x
- Brown, C.M., Holmes, R.B. & Currie, P.J. (2020) "A subadult individual of Styracosaurus albertensis (Ornithischia: Ceratopsidae) with comments on ontogeny and intraspecific variation in Styracosaurus and Centrosaurus" Vertebrate Anatomy Morphology Palaeontology, 8, 67-95. doi:10.18435/vamp29361
- Brown, C.M., Campione, N.E., Wilson Mantilla, G.P. & Evans, D.C. (2021) "Size-driven preservational and macroecological biases in the latest Maastrichtian terrestrial vertebrate assemblages of North America" Paleobiology. doi:10.1017/pab.2021.35
- Brown, C.M. & Henderson, D.M. (2015) "A new horned dinosaur reveals convergent evolution in cranial ornamentation in ceratopsidae" Current Biology, 25(12), 1641-1648. doi:10.1016/j.cub.2015.04.041
- Brown, R. W. (1962) "Paleocene flora of the Rocky Mountains and Great Plains" Geological Survey Professional Paper, 375, 1–119.
- BSM (online) http://www.thinktank.ac/page.asp?section=1074§ionTitle=Triceratops
- Campione, N.E. & Evans, D.C. (2012) "A universal scaling relationship between body mass and proximal limb bone dimensions in quadrupedal terrestrial tetrapods" BMC Biology, 10, 60 doi:10.1186/1741-7007-10-60
- Canoy Illies, M.M. & Fowler, D.W. (2020) "Triceratops with a kink: Co-ossification of five distal caudal vertebrae from the Hell Creek Formation of North Dakota, Cretaceous Research" 108, 104355. doi:10.1016/j.cretres.2019.104355
- Carpenter, K. (1979) "Vertebrate fauna of the Laramie Formation (Maestrichtian), Weld County, Colorado" Contributions to Geology, University of Wyoming, 17(1), 37-49.
- Carpenter, K. (2006) "Bison" alticornis and O.C. Marsh's early views on ceratopsians" [w:] Carpenter, K. (red.) "Horns and beaks: ceratopsian and ornithopod dinosaurs" Indiana University Press, 349-364.
- Carpenter, K. & Young, B. (2002) "Late Cretaceous dinosaurs from the Denver Basin, Colorado" [w:] Johnson, K.R., Raynolds, R.G. & Reynolds, M.L. (red.) "Paleontology and Stratigraphy of Laramide Strata in the Denver Basin, Pt. I." Rocky Mountain Geology, 37, 237-254. doi:10.2113/gsrocky.37.2.237
- Chinnery, B. (2004) "Morphometric analysis of evolutionary trends in the ceratopsian postcranial skeleton" Journal of Vertebrate Paleontology, 24(3), 591-609. doi:10.1671/0272-4634(2004)024[0591:MAOETI]2.0.CO;2
- Clark, B.R. (online 2015) https://www.facebook.com/photo.php?fbid=10153230237101775&set=p.10153230237101775&type=3
- Clemens, W.A. & Hartman, J.H. (2014) "From Tyrannosaurus rex to asteroid impact: Early studies (1901–1980) of the Hell Creek Formation in its type area" [w:] Wilson, G.P., Clemens, W.A., Horner, J.R. & Hartman, J.H. (red.) "Through the End of the Cretaceous in the Type Locality of the Hell Creek Formation in Montana and Adjacent Areas" The Geological Society of America Special Paper, 503, 1-87. doi:10.1130/2014.2503(01)
- CNM (online) https://nature.ca/collections-online
- CoBabe, E.A. & Fastovsky, D.E. (1987) "Ugrosaurus olsoni, a new ceratopsian (Reptilia: Ornithischia) from the Hell Creek Formation of eastern Montana" Journal of Paleontology, 61, 148-154.
- Colbert, E.H. (1962) "The weights of dinosaurs" American Museum Novitates, 2076, 1-16. http://hdl.handle.net/2246/3451
- CSC (online) http://www.csc.edu/sci/ebcmuseum/
- Currie, P.J., Holmes, R.B., Ryan, M.J., & Coy, C. (2016) "A juvenile chasmosaurine ceratopsid (Dinosauria, Ornithischia) from the Dinosaur Park Formation, Alberta, Canada" Journal of Vertebrate Paleontology, 36(2), e1048348. doi:10.1080/02724634.2015.1048348
- Curtice, B. (online 2021) https://www.facebook.com/groups/7636841670/posts/10157407078371671/?comment_id=10157407081251671&reply_comment_id=10157948827161671
- Cypress Hills-Grasslands Destination Area (online 2019) https://twitter.com/CHdestinationA/status/1177581285193052161
- Daley, A.C., & Budd, G.E. (2010) "New anomalocaridid appendages from the Burgess Shale, Canada" Palaeontology, 53(4), 721-738. doi:10.1111/j.1475-4983.2010.00955.x
- DMC (online a) http://www.dakotadino.com/Discovery.htm
- DMC (online b) http://www.dakotadino.com/Exhibits2.htm
- DeLuca, R. (online 2011) http://www.flickr.com/photos/ryno720/4667858397/
- CM (online) http://www.carnegiemnh.org/online/dinosaurs/triceratops.html
- Cope, E. D. (1889) "The Age of the Denver Formation" Science, 13(323), 290.
- D’Anastasio, R., Cilli, J., Bacchia, F., Fanti, F., Gobbo, G. & Capasso, L. (2021) "Combat Lesion in Triceratops: Histological and Chemical Diagnosis" PREPRINT (Version 1) Research Square doi:10.21203/rs.3.rs-1066311/v1
- D’Anastasio, R., Cilli, J., Bacchia, F., Fanti, F., Gobbo, G. & Capasso, L. (2022) "Histological and chemical diagnosis of a combat lesion in Triceratops" Scientific Reports, 12, 3941. doi:10.1038/s41598-022-08033-2
- Dalman, S.G., Lucas, S.G., Jasinki, S.E., Lichtig, A.J. & Dodson, P. (2021) "The oldest centrosaurine: a new ceratopsid dinosaur (Dinosauria: Ceratopsidae) from the Allison Member of the Menefee Formation (Upper Cretaceous, early Campanian), northwestern New Mexico, USA". PalZ, 95, 291-35. doi:10.1007/s12542-021-00555-w
- Davis, K. (online 2012) https://www.flickr.com/photos/planetdavis/8018147091/
- DePalma, R.A. (2010) "Geology, Taphonomy, and Paleoecology of a Unique Upper Cretaceous Bonebed Near the Cretaceous-Tertiary Boundary in South Dakota" praca magisterska, University of Kansas, Lawrence. http://hdl.handle.net/1808/6419
- Detrich, R. (online 2013) https://www.facebook.com/groups/7636841670/permalink/10151713127441671/
- Detrich, R. (online 2015a) https://twitter.com/kingfossil/status/609340901547053056
- Detrich, R. (online 2015b) https://www.facebook.com/groups/7636841670/permalink/10152908728516671/?comment_id=10152911072316671&comment_tracking=%7B%22tn%22%3A%22R%22%7D
- Detrich, R. (online 2015c) https://twitter.com/kingfossil/status/622967362568859648
- Detrich, R. (online 2017) https://twitter.com/kingfossil/status/819905291332501504
- Detrich, R. (online 2021) https://www.facebook.com/photo/?fbid=4301504179885121&set=p.4301504179885121
- Detrich, R. (online 2022) https://www.facebook.com/groups/7636841670/posts/10157971871446671/?comment_id=10157972198121671
- Dodson, P. (1996) "The Horned Dinosaurs" wyd. Princeton University, New Jersey, 1-346.
- Dodson, P. (2013) "Ceratopsia increase: history and trends" Canadian Journal of Earth Sciences, 50(3), 294-305. doi:10.1139/cjes-2012-0085
- Dodson, P., Coombs, W.P. Jr., Farlow, J.O. & Tatarinov, L.P. (1990) "Dinosaur Paleobiology" [w:] Weishampel, D.B., Dodson, P. & Osmólska, H. "The Dinosauria" University of California Press, 31-63.
- Dodson, P., Forster, C.A. & Sampson, S.D (2004) "Ceratopsidae" [w:] Weishampel, D.B., Dodson, P. & Osmólska, H. "The Dinosauria" University of California Press, 494-513.
- Drevermann, F. (1913) "Der Triceratops" Bericht über die Senckenbergische Naturforschende Gesellschaft in Frankfurt am Main, 44(2), 10-13. http://biodiversitylibrary.org/page/8843896
- Eberth, D.A. (1997) "Edmonton Group" [w:] Currie, P.J. & Padian. K. (red.) "Encyclopedia of Dinosaurs" Acaedemic Press/Elsevier, 199-204.
- Eberth, D.A., Evans, D.C., Brinkman, D., Therrien, F., Tanke, D.H.T. & Russell, L.S. (2013) "Dinosaur Biostratigraphy of the Edmonton Group (Upper Cretaceous), Alberta, Canada: Evidence for Climate Influence" Canadian Journal of Earth Sciences, 50(7), 701-726. doi:10.1139/cjes-2012-0185
- Eberth, D.A. & Kamo, S.L. (2020) "High-precision U–Pb CA–ID–TIMS dating and chronostratigraphy of the dinosaur-rich Horseshoe Canyon Formation (Upper Cretaceous, Campanian–Maastrichtian), Red Deer River valley, Alberta, Canada" Canadian Journal of Earth Sciences, 57 (10), 1220-1237. doi:10.1139/cjes-2019-0019
- EM (online 2012) http://eastendhistoricalmuseum.com/the-fossil-collection/
- EOL (online) http://eol.org/pages/757082/details
- Erickson, B.R. (1966) "Mounted skeleton of Triceratops prorsus in the Science Museum" Scientific Publications of the Science Museum of Minnesota, 1, 1-15.
- Estes, J.N. (online 2013) http://www.boston.com/community/moms/blogs/parent_buzz/2013/12/dinosaur_walkabout_cambridge_through_feb_2_2014.html
- Evans, D.C., Barrett, P.M. & Seymour, K.L. (2012) "Revised identification of a reported Iguanodon-grade ornithopod tooth from the Scollard Formation, Alberta, Canada" Cretaceous Research, 33(1), 11-14. doi:10.1016/j.cretres.2011.07.002
- Evans, D. (online 2014) https://www.facebook.com/photo.php?fbid=470111106460749&set=p.470111106460749&type=3&theater
- Evanson, T. (online 2015) https://www.flickr.com/photos/timevanson/27610627745
- Evenor, Z. (online 2014) https://www.flickr.com/photos/zachievenor/15413935239/
- Farke, A.A. (1997) "The distribution and taxonomy of Triceratops" [w:] Wolberg, D.L. Stump, E. & Rosenberg, G. (red.) "Dinofest International: Proceedings of a Symposium Held at Arizona State University. Philadelphia" Academy of Natural Sciences, 47-49.
- Farke, A.A. (2004) "Horn use in Triceratops (Dinosauria: Ceratopsidae): testing behavioral hypotheses using scale models" Palaeontologia Electronica, 7(1), 1-10.
- Farke, A.A. (online 2004) http://dml.cmnh.org/2004Apr/msg00544.html
- Farke, A.A. (online 2005) http://dml.cmnh.org/2005Apr/msg00273.html
- Farke, A.A. (2006) "Morphology and ontogeny of the cornual sinuses in chasmosaurine dinosaurs (Ornithischia: Ceratopsidae)" Journal of Paleontology, 80(4), 780-785. doi:10.1666/0022-3360(2006)80[780:MAOOTC]2.0.CO;2
- Farke, A.A. (2006) "Cranial osteology and phylogenetic relationships of the chasmosaurine ceratopsid Torosaurus latus" [w:] Carpenter, K. (red.) "Horns and beaks: ceratopsian and ornithopod dinosaurs" Bloomington, wyd. Indiana University Press, 235-257.
- Farke, A.A. (2008) "Function and Evolution of the Cranial Sinuses in Bovid Mammals and Ceratopsian Dinosaurs" dysertacja doktorska, Stony Book University, 1-213.
- Farke, A.A. (2010) "Evolution, homology, and function of the supracranial sinuses in ceratopsian dinosaurs" Journal of Vertebrate Palaeontology, 30(5), 1486-1500. doi:10.1080/02724634.2010.501436
- Farke, A.A. (2011) "Anatomy and Taxonomic Status of the Chasmosaurine Ceratopsid Nedoceratops hatcheri from the Upper Cretaceous Lance Formation of Wyoming, U.S.A." PLoS ONE 6(1), e16196. doi:10.1371/journal.pone.0016196
- Farke, A.A. (online 2012a) "Notes and Observations on Specimens of Torosaurus at the Yale Peabody Museum of Natural History" figshare doi:10.6084/m9.figshare.91333
- Farke, A.A (online 2012b) "Notes on Ceratopsians at the National Museum of Natural History" figshare doi:10.6084/m9.figshare.91349
- Farke, A.A. (online, 2012c) "Measurements of Ceratopsid Skulls" figshare doi:10.6084/m9.figshare.92648
- Farke, A.A. (online, 2012d) "Notes on Ceratopsians and Ankylosaurs at the Royal Ontario Museum" figshare doi:10.6084/m9.figshare.92066.v1
- Farke, A.A (online 2013) "Notes on Ceratopsians at the Canadian Museum of Nature" figshare doi:10.6084/m9.figshare.645336
- Farke, A.A. (2014) "Evaluating combat in ornithischian dinosaurs" Journal of Zoology, 292(4), 242-249. doi:10.1111/jzo.12111
- Farke, A.A (online 2016) https://www.facebook.com/groups/7636841670/posts/10153234344511671/?comment_id=10153234396341671
- Farke, A.A. (online 2018) https://www.facebook.com/groups/7636841670/posts/10155732033121671/
- Farke, A.A., Wolff, E.D.S. & Tanke, D.H. (2009) "Evidence of Combat in Triceratops" PLoS ONE 4(1), e4252. doi:10.1371/journal.pone.0004252
- Farke, A.A. & Alley, W. (2006) "An abnormal squamosal of Triceratops (Ornithischia: Ceratopsidae) from the Upper Cretaceous Hell Creek Formation of South Dakota" [w:] Lucas, S.G. & Sullivan, R.M. (red.) "Late Cretaceous vertebrates from the Western Interior" New Mexico Museum of Natural History and Science Bulletin, 35, 371-372.
- Ford, T., online - Nedoceratops http://www.paleofile.com/Dinosaurs/Ornithopods/Diceratops.asp
- Ford, T., online - Triceratops http://www.paleofile.com/Dinosaurs/Ornithopods/Triceratops.asp
- Ford, T., online - Triceratops sp. http://www.paleofile.com/Dinosaurs/Ornithopods/Triceratopssp.asp
- Ford, T., online - Torosaurus http://www.paleofile.com/Dinosaurs/Ornithopods/Torosaurus.asp
- Forster, C.A. (1990) "The cranial morphology and systematics of Triceratops, with a preliminary analysis of ceratopsian phylogeny" dysertacja doktorska, University of Pennsylvania, 1-227.
- Forster, C.A. (1993) "Taxonomic validity of the ceratopsid dinosaur Ugrosaurus olsoni (Cobabe and Fastovsky)" Journal of Paleontology, 67(2), 316-318.
- Forster, C.A. (1996a) "New Information on the Skull of Triceratops" Journal of Vertebrate Paleontology, 16 (2), 246-258. doi:10.1080/02724634.1996.10011312
- Forster, C.A. (1996b) "Species resolution in Triceratops: cladistic and morphometric approaches" Journal of Vertebrate Paleontology, 16 (2), 259-270. doi:10.1080/02724634.1996.10011313
- Forster, C.A., Sereno, P.C., Evans, T.W. & Rowe, T. (1993) "A complete skull of Chasmosaurus mariscalensis (Dinosauria, Ceratopsidae) from the Aguja Formation (Late Campanian) of west Texas" Journal of Vertebrate Paleontology, 13(2), 161-170. doi:10.1080/02724634.1993.10011498
- Fiorillo, A.R. & Tykoski, R.S. (2012) "A new Maastrichtian species of the centrosaurine ceratopsid Pachyrhinosaurus from the North Slope of Alaska" Acta Palaeontologica Polonica, 57(3), 561-573. doi:10.4202/app.2011.0033
- Fiorillo, A.R. & Tykoski, R.S. (2013) "An Immature Pachyrhinosaurus perotorum (Dinosauria: Ceratopsidae) Nasal Reveals Unexpected Complexity of Craniofacial Ontogeny and Integument in Pachyrhinosaurus" PLoS ONE, 8(6), e65802. doi:10.1371/journal.pone.0065802
- Fowler, D.W. (2010) "Anagenesis and long-term morphologic trends in Chasmosaurinae (Dinosauria: Ceratopsidae) revealed by a new high-resolution chronostratigraphic framework, ontogenetic analysis, and description of two new taxa" Journal of Vertebrate Paleontology 30(5, suplement), 91A. [abstrakt]
- Fowler, D.W. (online 2015) https://www.facebook.com/groups/7636841670/posts/10153133726856671/?comment_id=10153133749401671
- Fowler, D.W. (online 2016) "A new correlation of the Cretaceous formations of the Western Interior of the United States, I: Santonian-Maastrichtian formations and dinosaur biostratigraphy" PeerJ Preprints, 4, e2554v1 doi:10.7287/peerj.preprints.2554v1
- Fowler, D.W. (2017) "Revised geochronology, correlation, and dinosaur stratigraphic ranges of the Santonian-Maastrichtian (Late Cretaceous) formations of the Western Interior of North America" PLoS ONE, 12(11), e0188426. doi:10.1371/journal.pone.0188426
- Fowler, D.W. (online 2017) https://peerj.com/preprints/2554v1/#feedback-931
- Fowler, D.W. (online 2020) https://www.facebook.com/groups/7636841670/posts/10157312631446671/?comment_id=10157312934641671&reply_comment_id=10157313219431671
- Fowler, D.W., Scannella, J.B., & Horner, J.R (2011) "Reassessing ceratopsid diversity using unified frames of reference" Journal of Vertebrate Paleontology 31(5, suplement), 111. [abstrakt]
- Frederickson, J.A. (2012) "Cranial Deveopment of Centrosaurus apertus: understanding horn variation and evolution through an ontogenetic approach" Journal of Vertebrate Paleontology, 32(5, suplement), 97 [abstrakt].
- Frederickson, J.A. & Tumarkin-Deratzian, A.R. (2014) "Craniofacial ontogeny in Centrosaurus apertus" PeerJ, 2, e252. doi:10.7717/peerj.252
- Fujiwara, S.-i. (2009) "A Reevaluation of the manus structure in Triceratops (Ceratopsia: Ceratopsidae)". Journal of Vertebrate Paleontology, 29(4), 1136-1147. doi:10.1671/039.029.0406
- Fujiwara, S.-i. & Hutchinson, J.R. (2012) "Elbow joint adductor moment arm as an indicator of forelimb posture in extinct quadrupedal tetrapods" Proceedings of the Royal Society B: Biological Sciences, 279 (1738), 2561-2570. doi:10.1098/rspb.2012.0190
- Fujiwara, S.-i. & Takakuwa, Y. (2011) "A sub-adult growth stage indicated in the degree of suture co-ossification in Triceratops" Bulletin of the Gunma Museum of Natural History, 15: 1-17.
- Galton, P.M. & Sues, H.-D. (1983) "New data on pachycephalosaurid dinosaurs (Reptilia: Ornithischia) from North America" Canadian Journal of Earth Sciences, 20(3): 462-472.
- Garstka, W. & D.A. Burnham (1997) "Posture and stance of Triceratops: evidence of digitigrade manus and cantilever vertebral column" [w:] Wolberg, D.L., Stump, E. & Rosenberg, G.D. (red.) "Dinofest International Proceedings" Academy of Natural Sciences, Philadelphia, Pennsylvania: 385-391.
- "GetAwayTrike" (online 2014) http://blogs.yahoo.co.jp/rboz_05/33982954.html
- "GetAwayTrike" (online 2021) https://getawaytrike.hatenablog.com/entry/2021/08/17/164222
- Gilmore, C.W. (1906) "The mounted skeleton of Triceratops prorsus" Proceedings of the United States National Museum, 29, 433-435. http://biodiversitylibrary.org/page/15666173
- Gilmore, C. W., (1920) "A new restoration of Triceratops, with notes on the osteology of the genus" Proceedings of the United States National Museum, 55: 97-112.
- GLAHM (online 2017) http://www.huntsearch.gla.ac.uk/cgi-bin/foxweb/huntsearch/DetailedResults.fwx?collection=all&searchTerm=152339&mdaCode=GLAHM
- Glut, D.F. (1997) "Dinosaurs: The Encyclopedia" wyd. McFarland & Company
- Glut, D.F. (2002) "Dinosaurs: The Encyclopedia. Supplement 2" wyd. McFarland & Company
- Glut, D.F. (2003) "Dinosaurs: The Encyclopedia. Supplement 3" wyd. McFarland & Company
- Glut, D.F. (2006) "Dinosaurs: The Encyclopedia. Supplement 4" wyd. McFarland & Company
- Glut, D.F. (2008) "Dinosaurs: The Encyclopedia. Supplement 5" wyd. McFarland & Company
- Goodwin, M.B., Buchholtz, E.A. & Johnson, R.E. (1998) "Cranial anatomy and diagnosis of Stygimoloch spinifer (Ornithischia: Pachycephalosauria) with comments on cranial display structures in agonistic behavior" Journal of Vertebrate Paleontology, 18(2), 363-375.
- Goodwin, M. B., Clemens, W. A., Horner, J. R. & Padian, K. (2006) "The smallest known Triceratops skull: new observations on ceratopsid cranial anatomy and ontogeny" Journal of Vertebrate Paleontology, 26(1), 103-112. doi:10.1671/0272-4634(2006)26[103:TSKTSN]2.0.CO;2
- Goodwin, M.B. & Horner, J.R. (2001) "How Triceratops got its horns: New information from a growth series on cranial morphology and ontogeny" Journal of Vertebrate Paleontology, 21(3 - supplement), 56A [abstrakt]
- Goodwin, M.B. & Horner, J.R. (2010) "Historical Collecting Bias and the Fossil Record of Triceratops in Montana" [w:] Ryan, M.J., Chinnery-Allgeier, B.J. & Eberth, D.A. (red.) "New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium" Indiana University Press, Bloomington, 551-563.
- Goodwin, M.B. & Horner, J.R. (2014) "Cranial morphology of a juvenile Triceratops skull from the Hell Creek Formation, McCone County, Montana, with comments on the fossil record of ontogenetically younger skulls" [w:] Wilson, G.P., Clemens, W.A., Horner, J.R. & Hartman, J.H. (red.) "Through the End of the Cretaceous in the Type Locality of the Hell Creek Formation in Montana and Adjacent Areas" The Geological Society of America Special Paper, 503, 333-348. doi:10.1130/2014.2503(13)
- Goussard, F. (2006) "The skull of Triceratops in the palaeontology gallery, Muséum national d’Histoire naturelle, Paris" Geodiversitas, 28(3), 467-476.
- Graham, M.R. (online 2016) https://www.facebook.com/groups/7636841670/permalink/10153200291611671/
- gramunion.com (online) http://www.gramunion.com/triceratops-the-fighter.tumblr.com/148157458465
- GRM (online) http://www.thegrandrivermuseum.com/new-gallery-5/
- GRM (online 2012) http://grandrivermuseum.blogspot.com/2012/05/grand-river-museum-is-on-highway-12-in.html
- GRM (online 2013) http://www.grandrivermuseum.org/dinosaurs.htm
- Hall, L. (online 2016) https://www.facebook.com/groups/7636841670/permalink/10153699990851671/
- Happ, J. (2008) "An analysis of predator-prey behavior in a head-to-head encounter between Tyrannosaurus rex and Triceratops" [w:] Larson, P. & Carpenter, K. (red.) "Tyrannosaurus rex: the tyrant king" Indiana University Press, 354-368.
- Happ, J.W. (2010) "New Evidence Regarding the Structure and Function of the Horns in Triceratops (Dinosauria: Ceratopsidae[w:] Ryan, M.J., Chinnery-Allgeier, B.J. & Eberth, D.A. (red.) "New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium" Indiana University Press, Bloomington, 271-281.
- Happ, J.W. & Morrow C.M. (1996) "Separation of Triceratops (Dinosauria: Ceratopsidae) into two allopatric species by cranial morphology" Journal of Vertebrate Paleontology 16 (3, suplement), Abstracts of Papers, Fifty-Sixth Annual Meeting, 40A. [abstrakt]
- Happ, J.W. & Morrow, C.M. (2000) "Evidence of soft tissue associated wiith nasal and supraorbital horn cores, rostral and exoccipital of Triceratops" Journal of Vertebrate Paleontology, 20(3 - suplement), 47A. [abstrakt]
- Harper, H. (online 2014) https://www.flickr.com/photos/sethward/14015714986
- Hatcher, J.B. (1905) "Two new Ceratopsia from the Laramie of Converse County, Wyoming" American Journal of Science, Series 4, 120, 413-419. http://biodiversitylibrary.org/page/40245265
- Hatcher, J.B., Marsh, O.C., & Lull, R.S. (1907) "The Ceratopsia" US Geological Survey Monograph, 49, 1-300.
- Henderson, M. (online 2012) http://gizzardstone.com/gizzard-stones/
- Hendrickx, C. & Bell, P.R. (2021) "The scaly skin of the abelisaurid Carnotaurus sastrei (Theropoda: Ceratosauria) from the Upper Cretaceous of Patagonia" Cretaceous Research, 128, 104994. doi:10.1016/j.cretres.2021.104994
- Heritage Auctions (online 2018) https://fineart.ha.com/itm/fossils/dinosauria/dinosaur-skullceratopsianlate-cretaceoushell-creek-formationharding-county-south/a/5389-72115.s
- Hieronymus, T.L., Witmer, L.M., Tanke, D.H. & Currie, P.J. (2009) "The facial integument of centrosaurine ceratopsids: morphological and histological correlates of novel skin structures" Anatomical Record, 292(9), 1370-1396. doi:10.1002/ar.20985
- Hicks, J.F., Johnson, K.R., Obradovich, J.D., Tauxe, L. & Clark, D. (2002) "Magnetostratigraphy and geochronology of the Hell Creek and basal Fort Union Formations of southwestern North Dakota and a recalibration of the age of the Cretaceous-Tertiary boundary" [w:] Hartman, J.H., Johnson, K.R. & Nichols, D.J. "The Hell Creek Formation and the Cretaceous-Tertiary boundary in the northern Great Plains: An Integrated continental record of the end of the Cretaceous" Geological Society of America Special Paper, 361, 35-55.
- Hoganson, J.W. (online 2006) https://www.dmr.nd.gov/ndfossil/Exhibits/Corridor%20of%20Time%20Exhibit%20030620.html
- Holland, F.D. (1997) "A North Dakota Triceratops skull" Rocky Mountain Geology, 32(1), 51-60.
- Holliday, C.M. (2009) "New insights into dinosaur jaw muscle anatomy" The Anatomical Record, 292(9), 1246-1265. doi:10.1002/ar.20982
- Horner, J.R. & Goodwin, M.B. (2005) "A new Triceratops cranial growth series" Journal of Vertebrate Paleontology, 25 (3 - suplement), 71A-72A. [abstrakt]
- Horner, J.R. & Goodwin, M.B. (2006) "Major cranial changes during Triceratops ontogeny" Proceedings of the Royal Society B: Biological Sciences, 273(1602), 2757-2761. doi:10.1098/rspb.2006.3643
- Horner, J.R. & Goodwin, M.B. (2008) "Ontogeny of cranial epi-ossifications in Triceratops" Journal of Vertebrate Paleontology, 28(1), 134-144. doi:10.1671/0272-4634(2008)28[134:OOCEIT]2.0.CO;2
- Horner, J.R. & Goodwin, M.B. (2009) "Extreme Cranial Ontogeny in the Upper Cretaceous Dinosaur Pachycephalosaurus" PLoS ONE, 4(10), e7626. doi:10.1371/journal.pone.0007626
- Horner, J.R. & Lamm, E.T. (2011) "Ontogeny of the parietal frill of Triceratops: a preliminary histological analysis" Comptes Rendus Palevol, 10(5-6), 439-452. doi:10.1016/j.crpv.2011.04.006
- Horner, J.R. & Marshall, C. (2002) "Keratinous covered dinosaur skulls" Journal of Vertebrate Paleontology, 22(3 - suplement), 67A. [abstrakt]
- Horner, J.R., Goodwin, M.B. & Myhrvold, N. (2011) "Dinosaur Census Reveals Abundant Tyrannosaurus and Rare Ontogenetic Stages in the Upper Cretaceous Hell Creek Formation (Maastrichtian), Montana, USA" PLoS ONE, 6(2), e16574. doi:10.1371/journal.pone.0016574
- Howie, C. (online 2010) https://commons.wikimedia.org/wiki/File:TCMI_Triceratops_restoration.tif
- Hudson, G. (online 2008) http://www.flickr.com/photos/grum/2819782317/
- Huigen, L. (online 2013) https://science.naturalis.nl/en/about-us/news/trexexpeditie/rare-skeleton-of-well-known-dinosaur/
- Hunt-Foster, R.K. (online 2009) http://paleochick.blogspot.com/2009/03/first-triceratops-bonebed.html
- Hurt, S. (online) "Triceratops Cranial Ontogeny: How the skull of Triceratops developed from birth to adulthood" http://www.nhm.org/site/sites/default/files/dinosaur_institute/pdf/NHM_DI_Sean_Hurt_Triceratops_Ontogeny.pdf
- iknowdino.com (online 2015) http://iknowdino.com/hadrosaurus-foulkii-at-the-academy-of-natural-sciences-of-drexel-university/
- International Chronostratigraphic Chart, International Commission on Stratigraphy (2013) http://www.stratigraphy.org/ICSchart/ChronostratChart2013-01.pdf
- Johnson, T. (online 2014a) https://www.facebook.com/groups/7636841670/permalink/10151951512801671/
- Johnson, T. (online 2014b) https://www.facebook.com/groups/7636841670/permalink/10152064066626671/
- Johnson, T. (online 2014c) https://www.facebook.com/groups/7636841670/permalink/10152181169311671/?comment_id=10152188163546671&comment_tracking=%7B%22tn%22%3A%22R1%22%7D
- Johnson, T. (online 2014d) https://www.facebook.com/groups/7636841670/permalink/10152309849406671/
- Johnson, T. (online 2022) https://www.facebook.com/groups/7636841670/posts/10158202705506671/
- Jones, J. (online 2016) http://listen.sdpb.org/post/quadruple-triceratops-dig-gives-insight-dinosaur-behavior
- Kato, H. (online 2019) https://www.facebook.com/media/set/?set=oa.10156331767691671&type=3
- Kato, H. (online 2021) https://www.facebook.com/groups/7636841670/posts/10158109092356671/
- "KCJ44" (online 2014) https://www.tripadvisor.com/Attraction_Review-g32066-d117264-Reviews-Lawrence_Hall_of_Science-Berkeley_California.html#photos;geo=32066&detail=117264&ff=96367781
- Keenan, S.W. & Scannella, J.B. (2014) "Paleobiological implications of a Triceratops bonebed from the Hell Creek Formation, Garfield County, northeastern Montana" [w:] Wilson, G.P., Clemens, W.A., Horner, J.R. & Hartman, J.H. (red.) "Through the End of the Cretaceous in the Type Locality of the Hell Creek Formation in Montana and Adjacent Areas" The Geological Society of America Special Paper, 503, 349-364. doi:10.1130/2014.2503(14)
- Krauss, D.A., Pezon, A., Nguyen, P., Salame, I. & Rywkin, S.B. (2010) "Evolutionary Interactions between Horn and Frill Morphology in Chasmosaurine Ceratopsians" [w:] Ryan, M.J., Chinnery-Allgeier, B.J. & Eberth, D.A. (red.) "New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium" Indiana University Press, Bloomington, 282-292.
- Król Padilha, P. (online 2008) http://www.ikessauro.com/2008/02/tricertops.html
- LACM (online) https://collections.nhm.org/dinosaur-institute/
- Lampkins, L. (online 2017) https://www.facebook.com/groups/7636841670/permalink/10154317167331671/
- Larson, N. (online 2015a) https://www.facebook.com/photo.php?fbid=10153275552169902&set=a.10151324105014902.470828.551134901&type=3
- Larson, N. (online 2015b) https://www.facebook.com/larsonpaleontology/posts/10153275903354902
- Larson, N. (online 2015c) https://www.facebook.com/larsonpaleontology/posts/10153276442569902
- Larson, N. (online 2015d) https://www.facebook.com/bob.detrich/posts/860428267326080?comment_id=860533150648925&comment_tracking=%7B%22tn%22%3A%22R0%22%7D
- Larson, P. (online 2015) https://twitter.com/petelarsontrex/status/604611242553188352?lang=en
- Larson, P., Evans, D. & Ott, C. (2004) "Triceresies - a bold new look at Triceratops" Journal of Vertebrate Paleontology, 24(3 - suplement), 82A. [abstrakt]
- Larson, P.L., Larson, M., Ott, C., Bakker, R. (2007) "Skinning a Triceratops" Journal of Vertebrate Paleontology, 27(3 - supplement), 102A [abstrakt]
- Larson Paleontology Unlimited (online 2014a) https://www.facebook.com/LarsonPaleontologyUnlimited/photos/pcb.805296396180511/805295836180567/?type=3&theater
- Larson Paleontology Unlimited (online 2014b) https://www.facebook.com/LarsonPaleontologyUnlimited/photos/a.701724323204386/815920748451409/
- Larson Paleontology Unlimited (online 2015a) https://www.facebook.com/LarsonPaleontologyUnlimited/posts/973094982733984
- Larson Paleontology Unlimited (online 2015b) https://www.facebook.com/LarsonPaleontologyUnlimited/photos/pcb.1007221515987997/1007219539321528/?type=3&theater
- LASM (online 2017) http://www.LASM.org/play-learn/hands-on-galleries/solar-system
- Leap, C. (online 2012a) http://blog.hmns.org/2012/05/introducing-lane-a-mummified-triceratops-with-a-new-address-at-5555-hermann-park-dr/
- Leap, C. (online 2012b) http://blog.hmns.org/2012/05/the-making-of-a-moniker-how-lane-got-his-name-and-a-wyoming-teenager-got-a-spot-in-the-history-books/
- Lee, Y.M. (online 2014) https://www.facebook.com/groups/7636841670/permalink/10152464504966671/?comment_id=10152465189006671&comment_tracking=%7B%22tn%22%3A%22R3%22%7D
- Lehman, T. M. (1990) "The ceratopsian subfamily chasmosaurinae: sexual dimorphism and systematics" [w:] Carpenter, K. & Currie, P.J. (red.) "Dinosaur Systematics: Approaches and Perspectives" wyd. Cambridge University, New York, 211-229.
- Lehman, T.M. (1996) "A horned dinosaur from the El Picacho Formation of West Texas, and review of ceratopsian dinosaurs from the American Southwest" Journal of Paleontology, 70(3), 494-508.
- Lehman, T.M. (1998) "A gigantic skull and skeleton of the horned dinosaur Pentaceratops sternbergi from New Mexico" Journal of Paleontology, 72(5), 894-906.
- Lehman, T.M. (2006) "Growth and population age structure in the horned dinosaur Chasmosaurus" [w:] Carpenter, K. (red.) "Horns and beaks: ceratopsian and ornithopod dinosaurs" Bloomington, wyd. Indiana University Press, 259-317.
- Lockley, M.G. & Hunt, A.P. (1995) "Ceratopsid tracks and associated ichnofauna from the Laramie Formation (Upper Cretaceous: Maastrichtian) of Colorado" Journal of Vertebrate Paleontology, 15(3), 592-614.
- Longrich, N.R. (2010) "Mojoceratops perifania, A New Chasmosaurine Ceratopsid from the Late Campanian of Western Canada" Journal of Paleontology, 84 (4), 681-694.
- Longrich, N. R. (2011) "Titanoceratops ouranos, a giant horned dinosaur from the late Campanian of New Mexico" Cretaceous Research, 32(3), 264-276. doi:10.1016/j.cretres.2010.12.007
- Longrich, N.R. (2014) "The horned dinosaurs Pentaceratops and Kosmoceratops from the upper Campanian of Alberta and implications for dinosaur biogeography" Cretaceous Research, 51, 292-308. doi:10.1016/j.cretres.2014.06.011
- Longrich, N. (online 2015) "Systematics of Chasmosaurus - new information from the Peabody Museum skull, and the use of phylogenetic analysis for dinosaur alpha taxonomy" [version 1; referees: 1 not approved] F1000Research, 4, 1468. doi:10.12688/f1000research.7573.1
- Longrich, N.R. & Field, D.J. (2012) "Torosaurus Is Not Triceratops: Ontogeny in Chasmosaurine Ceratopsids as a Case Study in Dinosaur Taxonomy" PLoS ONE 7(2), e32623. doi:10.1371/journal.pone.0032623
- Longrich, N.R., Horner, J.R., Erickson, G.M. & Currie, P.J. (2010) "Cannibalism in Tyrannosaurus rex" PLoS ONE 5(10), e13419. doi:10.1371/journal.pone.0013419
- Lull, R.S. (1903) "Skull of Triceratops serratus" Bulletin of the American Museum of Natural History, 19, 685-695. http://hdl.handle.net/2246/1509
- Lull, R.S. (1905) "Restoration of the horned dinosaur Diceratops" American Journal of Science, Series 4, 4, 420-422.
- Lull, R.S. (1915) "The mammals and horned dinosaurs of the Lance Formation of Niobrara County, Wyoming" American Journal of Science, series 4, 40, 319-348. http://biodiversitylibrary.org/page/40389337
- Lull, R.S. (1933) "A revision of the Ceratopsia, or horned dinosaurs" Memoirs of the Peabody Museum of Natural History, 3, 1-175.
- Lull, R.S. (1934) "Skull of Triceratops flabellatus recently mounted at Yale" American Journal of Science, series 5, 28, 439-442. doi:10.2475/ajs.s5-28.168.439
- Lund, E.K., Sampson, S.D., & Loewen, M.A. (2016) "Nasutoceratops titusi (Ornithischia, Ceratopsidae), a basal centrosaurine ceratopsid from the Kaiparowits Formation, southern Utah" Journal of Vertebrate Paleontology, 36(2), e1054936. doi:10.1080/02724634.2015.1054936
- Lund, S.P., Hartman, J.H., & Banerjee, S.K. (2002) "Magnetostratigraphy of interfingering upper Cretaceous-Paleocene marine and continental strata of the Williston Basin, North Dakota and Montana" [w:] Hartman, J.H., Johnson, K.R. & Nichols, D.J. "The Hell Creek Formation and the Cretaceous-Tertiary boundary in the northern Great Plains: An Integrated continental record of the end of the Cretaceous" Geological Society of America Special Paper, 361, 57-74.
- Lydekker, R. (1893) "Some Recent Restorations of Dinosaurs" Nature, 48(1239), 302-304. doi:10.1038/048302a0
- Lyson, T.R. & Longrich, N.R. (2011) "Spatial niche partitioning in dinosaurs from the latest cretaceous (Maastrichtian) of North America" Proceedings of the Royal Society B: Biological Sciences, 278(1709), 1158-1164. doi:10.1098/rspb.2010.1444
- Mabbett, A. (online 2015) https://en.wikipedia.org/w/index.php?title=Thinktank%2C_Birmingham_Science_Museum&type=revision&diff=649977796&oldid=645890473#cite_note-16
- Maidment, S.C.R. & Barrett, P.M. (2011) "A new specimen of Chasmosaurus belli (Ornithischia: Ceratopsidae), a revision of the genus, and the utility of postcrania in the taxonomy and systematics of ceratopsid dinosaurs" Zootaxa, 2963, 1-47.
- Maidment, S.C.R., Linton, D.H., Upchurch, P. & Barrett, P.M. (2012) "Limb-Bone Scaling Indicates Diverse Stance and Gait in Quadrupedal Ornithischian Dinosaurs" PLoS ONE, 7(5), e36904. doi:10.1371/journal.pone.0036904
- Maiorino, L, Farke, A.A., Piras, P., Ryan, M.J., Terris, K.M. & Kotsakis, T. (2013a) "The evolution of squamosal shape in ceratopsid dinosaurs (Dinosauria, Ornithischia)" Journal of Vertebrate Paleontology, 33(6), 1385-1393, doi:10.1080/02724634.2013.779922
- Maiorino, L., Farke, A.A., Kotsakis, T. & Piras, P. (2013b) "Is Torosaurus Triceratops? Geometric Morphometric Evidence of Late Maastrichtian Ceratopsid Dinosaurs" PLoS ONE, 8(11), e81608. doi:10.1371/journal.pone.0081608
- Maiorino, L., Farke, A.A., Kotsakis, T., Teresi, L. & Piras, P. (2015) "Variation in the shape and mechanical performance of the lower jaws in ceratopsid dinosaurs (Ornithischia, Ceratopsia)" Journal of anatomy, 227(5), 631-646. doi:10.1111/joa.12374
- Maiorino, L., Farke, A.A., Kotsakis, T. & Piras, P. (2017) "Macroevolutionary patterns in cranial and lower jaw shape of ceratopsian dinosaurs (Dinosauria, Ornithischia): phylogeny, morphological integration, and evolutionary rates" Evolutionary Ecology Research, 18, 123-167.
- Mallison, H. (online 2013) http://dinosaurpalaeo.wordpress.com/2013/12/14/dinosaur-mounts-at-the-nhm-l-a-triceratops/
- Mallon, J.C., Holmes, R., Eberth, D.A., Ryan, M.J. & Anderson, J.S. (2011) "Variation in the skull of Anchiceratops (Dinosauria, Ceratopsidae) from the Horseshoe Canyon Formation (Upper Cretaceous) of Alberta" Journal of Vertebrate Paleontology, 31(5), 1047-1071. doi:10.1080/02724634.2011.601484
- Mallon, J.C., Holmes, R., Anderson, J.S., Farke, A.A. & Evans, D.C. (2014a) "New Information on the Rare Horned Dinosaur Arrhinoceratops brachyops (Ornithischia: Ceratopsidae) from the Upper Cretaceous of Alberta, Canada" Canadian Journal of Earth Sciences. doi:10.1139/cjes-2014-0028
- Mallon J.C., Ott, C.J., Larson, P.L., Iuliano, E.M. & Evans, D.C. (2016) "Spiclypeus shipporum gen. et sp. nov., a Boldly Audacious New Chasmosaurine Ceratopsid (Dinosauria: Ornithischia) from the Judith River Formation (Upper Cretaceous: Campanian) of Montana, USA" PLoS ONE, 11(5), e0154218. doi:10.1371/journal.pone.0154218
- Maltese, A. (online 2021a) https://www.facebook.com/groups/7636841670/posts/10157918641116671/
- Maltese, A. (online 2021b) https://www.facebook.com/photo/?fbid=6347855891921931&set=p.6347855891921931
- Manning, P.L. & Egerton, V.M. (2014) "Taphonomy of a multi-taxa bone-bed in the Hell Creek Formation (Late Cretaceous) of South Dakota (USA)" [w:] "Proceedings of the Tenth Conference on Fossil Resources" Rapid City, 2014, 51-53. [abstrakt]
- Marsh, O.C. (1887) "Notice of some new fossil mammals" American Journal of Science, series 3, 34, 323-331.
- Marsh, O.C. (1889a) "Notice of new American dinosaurs" American Journal of Science, series 3, 37, 331-336.
- Marsh, O.C. (1889b) "Notice of gigantic horned Dinosauria from the Cretaceous" American Journal of Science, series 3, 38, 173-175.
- Marsh, O.C. (1889c) "The Skull of the Gigantic Ceratopsidae" American Journal of Science, series 3, 38, 500-506.
- Marsh, O.C. (1890a) "Description of new dinosaurian reptiles" American Journal of Science, series 3, 39: 81-86.
- Marsh, O.C. (1890b) "Additional characters of the Ceratopsidae, with notice of new Cretaceous dinosaurs" American Journal of Science, series 3, 39, 418-426.
- Marsh, O.C. (1891a) "Restoration of Triceratops" American Journal of Science, series 3, 41, 339-342.
- Marsh, O.C. (1891b) "Notice of new vertebrate fossils" American Journal of Science, series 3, 42, 265-269.
- Marsh, O.C. (1898) "New species of Ceratopsia" American Journal of Science, series 4, 6, 92.
- Matthew, W.D. (1915) "Dinosaurs with special reference to the American museum collections" American Museum of Natural History, New York.
- Mathews, J.C., Brusatte, S.L., Williams, S.A. & Henderson, M.D. (2009) "The first Triceratops bonebed and its implications for gregarious behavior" Journal of Vertebrate Paleontology, 29(1), 286-290. doi:10.1080/02724634.2009.10010382
- McDonald, A.T. (2011) "A Subadult Specimen of Rubeosaurus ovatus (Dinosauria: Ceratopsidae), with Observations on Other Ceratopsids from the Two Medicine Formation" PLoS ONE, 6(8), e22710. doi:10.1371/journal.pone.0022710
- McDonald, A. (online 2018a) https://canadianmuseumofnature.wordpress.com/2018/05/16/is-the-worlds-largest-triceratops-skull-sitting-in-our-collection/
- McDonald, A. (online 2018b) https://canadianmuseumofnature.wordpress.com/2018/12/12/triceratops-skull
- McIntosh, J.S. (1981) "Annotated catalogue of the Dinosaurs (Reptilia, Archosauria) in the Collections of Carnegie Museum of Natural Histor" Bulletin of the Carnegie Museum of Natural History, 18, 1-67.
- McLean, A. (online 2011) http://demitrivladmaximov.deviantart.com/art/Triceratops-251536156
- Melvin, J. (online 2016) http://newsstand.clemson.edu/mediarelations/to-brighten-its-future-bob-campbell-geology-museum-turns-to-a-dinosaur-from-the-past/
- Miller, B. (online 2013) https://dinosours.wordpress.com/2013/05/25/a-triceratops-for-lawrence/
- Milner, A.C. & Barrett, P.M. (2016) "Smith Woodward's contribution on fossil tetrapods" [w:] Johanson, Z., Barrett, P.M., Richter, M. & Smith, M. (red.) "Arthur Smith Woodward: His Life and Influence on Modern Vertebrate Palaeontology" Geological Society, London, Special Publications, 430, 289-309. doi:10.1144/SP430.13
- Milner, G. (online 2014) https://commons.wikimedia.org/wiki/File:Triceratops_Skeleton_-_National_Museum_of_Natural_History_(14611729991)a.jpg
- Molina Pérez, R. (online 2019) https://www.facebook.com/groups/7636841670/posts/10156245885801671/?comment_id=10156245896701671&reply_comment_id=10156245917471671
- Morales, M. & Campbell, C. (2006) "Variations from adult morphology in a subadult skull of the dinosaur Triceratops from the Hell Creek Formation of Montana" Journal of Vertebrate Paleontology, 26(3 - suplement), 103A. [abstrakt]
- MOR (online 2012) https://www.facebook.com/photo.php?fbid=532122583471080&set=a.532118976804774.102134976.230141317002543&type=3&theater
- MOR (online 2013) https://www.facebook.com/photo.php?fbid=635137613169576
- MOR (online 2014) https://www.facebook.com/photo.php?fbid=843600948989907
- MOR (online 2018) https://twitter.com/MORPaleo/status/972207143406288896
- MOR (online 2019a) https://www.facebook.com/mormsu/photos/a.96245075457/10155929143650458/
- MOR (online 2019b) https://www.facebook.com/mormsu/photos/pcb.10155970192725458/10155970192675458
- MOR (online 2020) https://www.facebook.com/mormsu/photos/pcb.10157240079930458/10157240079825458/
- MOR (online) http://muse.museum.montana.edu/paleodb_pub
- MOS (online) http://www.mos.org/exhibits/triceratops-cliff
- National Science Foundation (online 2010) http://www.nsf.gov/news/news_images.jsp?cntn_id=116480&org=NSF
- MWC (online 2011a) https://www.facebook.com/media/set/?set=a.258015607590946.60535.171256439600197&type=3
- MWC (online 2011b) https://scontent-b-lhr.xx.fbcdn.net/hphotos-frc3/382592_265571190168721_2047059675_n.jpg
- NBC (online) https://www.google.com/culturalinstitute/beta/asset/triceratops/6AFs4vcEmLIwdg?hl=en
- North Dakota Geological Survey (online) https://www.dmr.nd.gov/ndfossil/paleoregs/
- O'Connell, M.B. (online 2015) http://rapidcityjournal.com/news/local/dinosaur-skull-found-in-buffalo-likely-a-new-species/article_b33d3375-b74c-5941-aa0b-281ad59b430a.html
- Ogg, J.G. (2012) "Geomagnetic Polarity Time Scale" [w:] Gradstein, F.M., Ogg, J.G., Schmitz, M.D. & Ogg, G.M. (red.) "The Geologic Time Scale 2012" Elsevier, 85-113.
- Olshevsky, G. (online 2001) http://dml.cmnh.org/2001Aug/msg00241.html
- "Oregon Hiker" (online 2007a) http://www.flickr.com/photos/oregonhiker/935433255
- "Oregon Hiker" (online 2007b) http://www.flickr.com/photos/54767520@N00/935433213
- Orlowski, A. (online 2012) http://rapidcityjournal.com/news/fossil-hunter-says-triceratops-head-may-be-biggest-ever/article_f28b75eb-60ae-5c74-bae3-54a778dccb91.html
- Osborn, H.F. (1933) "Mounted skeleton of Triceratops elatus" American Museum Novitates, 654, 1-14. http://hdl.handle.net/2246/2066
- Ostrom, J. H. & Wellnhofer, P. (1986) "The Munich specimen of Triceratops with a revision of the genus" Zitteliana, 14: 111-158.
- Ott, C.J. & Larson, P.L. (2010) "A new, small ceratopsian dinosaur from the latest Cretaceous Hell Creek Formation, northwest South Dakota, United States: a preliminary description" [w:] Ryan, M.J., Chinnery, B.J. & Eberth, D.A. (red.) "New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium" wyd. Indiana University, Bloomington, 203-218.
- Paleobiology Database http://paleodb.org/?a=displayReference&reference_no=24896
- Paul, G.S. (1997) "Dinosaur models: the good, the bad, and using them to estimate the mass of dinosaurs" [w:] Wolberg, D.L., Stump, E. & Rosenberg, G.D. (red.) "Dinofest International Proceedings" Academy of Natural Sciences, Philadelphia, Pennsylvania, 129-154.
- Paul, G.S. (2010) "The Princeton Field Guide to Dinosaurs" wyd. Princeton University - Princeton i Oxford
- Paul, G.S. (online 2010) "Mass estimate table" http://www.gsdino.com/data.html
- Pearson, D.A., Schaefer, T., Johnson, K.R., Nichols, D.J., & Hunter, J.P., (2002) "Vertebrate biostratigraphy of the Hell Creek Formation in southwestern North Dakota and northwestern South Dakota" [w:] Hartman, J.H., Johnson, K.R. & Nichols, D.J. "The Hell Creek Formation and the Cretaceous-Tertiary boundary in the northern Great Plains: An Integrated continental record of the end of the Cretaceous" Geological Society of America Special Paper, 361, 145-167.
- Pence, L. (online 2013) https://drpence.wordpress.com/2013/08/07/dinosaurs
- Penkalski, P. & Skulan, J. (2010) "An unusual ceratopsid quarry from the Hell Creek Formation of Montana" Society of Vertebrate Paleontology Seventienth Anniversary Meeting Program and Abstracts Book, 145A. [abstrakt]
- Peterson, L. (online) https://plus.google.com/photos/photo/103422756128678618674/6433509363210096962
- Phillips, B. (online 2016) https://www.facebook.com/permalink.php?story_fbid=1039397019470212&id=102882843121639
- Porras-Múzquiz, H.G. & Lehman, T.M. (2011) "A ceratopsian horncore from the Olmos Formation (early Maastrichtian) near Múzquiz, Mexico" Revista Mexicana de Ciencias Geológicas, 28(2), 262-266.
- Prieto-Márquez, A., Garcia-Porta. J., Joshi, S.H., Norell, M.A. & Makovicky, P.J. (2020) "Modularity and heterochrony in the evolution of the ceratopsian dinosaur frill" Ecology and Evolution, 10, 6288-6309. doi:10.1002/ece3.6361
- Rainforth, E.C. (2003) "Revision and re-evaluation of the Early Jurassic dinosaurian ichnogenus Otozoum" Palaeontology, 46(4), 803-838. doi:10.1111/1475-4983.00320
- Roberts, E.K. (2019) "Description of a Juvenile Triceratops Skull and Lower Jaw and Comparison of Juvenile Ceratopsian Characteristics" praca magisterska, Emporia State University. 1-66.
- Rogers, K. & Rafferty, J. (online 2010) http://www.britannica.com/blogs/2010/09/torosaurus-is-triceratops-5-questions-for-paleontologist-john-scannella/
- Rolfes, M. (online 2017) https://www.facebook.com/photo.php?fbid=459100661122689&set=pcb.10154645031401671&type=3&theater
- Russell, D.A. & Manabe, M. (2002) "Synopsis of the Hell Creek (uppermost Cretaceous) dinosaur assemblage" [w:] Hartman, J.H., Johnson, K.R. & Nichols, D.J. "The Hell Creek Formation and the Cretaceous-Tertiary boundary in the northern Great Plains: An Integrated continental record of the end of the Cretaceous" Geological Society of America Special Paper, 361, 169-176.
- Ryan, M.J. (2003) "Taxonomy, systematics and evolution of Centrosaurine Ceratopsids of the Campanian western interior basin of North America" dysertacja doktorska, University of Calgary, Calgary, Alta, 1-578.
- Ryan, M.J. (2007) "A new basal centrosaurine ceratopsid from the Oldman Formation, southeastern Alberta" Journal of Paleontology, 81(2), 376-396. doi:10.1666/0022-3360(2007)81[376:ANBCCF]2.0.CO;2
- Ryan, M.J. & Russell, A.P. (2005) "A new centrosaurine ceratopsid from the Oldman Formation of Alberta and its implications for centrosaurine taxonomy and systematics" Canadian Journal of Earth Sciences, 42 (7), 1369-1387. doi:10.1139/e05-029
- Sakagami, R. & Kawabe, S. (2020) "Endocranial anatomy of the ceratopsid dinosaur Triceratops and interpretations of sensory and motor function" PeerJ, 8, e9888. doi:10.7717/peerj.9888
- Sampson, S.D. (1995) "Two new horned dinosaurs from the Upper Cretaceous Two Medicine Formation of Montana; with a phylogenetic analysis of the Centrosaurinae (Ornithischia: Ceratopsidae)" Journal of Vertebrate Paleontology, 15(4), 743-760.
- Sampson, S.D. & Loewen M.A. (2010) "Unraveling a Radiation: A Review of the Diversity, Stratigraphic Distribution, Biogeography, and Evolution of Horned Dinosaurs (Ornithischia: Ceratopsidae)" [w:] Ryan, M.J., Chinnery-Allgeier, B.J. & Eberth, D.A. (red.) "New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium" Indiana University Press, Bloomington, 405-427.
- Sampson, S.D., Ryan, M.J. & Tanke, D.H. (1997) "Craniofacial ontogeny in centrosaurine dinosaurs (Ornithischia: Ceratopsidae): taxonomic and behavioral implications" Zoological Journal of the Linnean Society, 121(3), 293-337.
- Sampson, S.D., Loewen, M.A., Farke, A.A., Roberts, E.M., Forster, C.A., Smith, J.A. & Titus, A.L. (2010) "New Horned Dinosaurs from Utah Provide Evidence for Intracontinental Dinosaur Endemism" PLoS ONE, 5(9), e12292. doi:10.1371/journal.pone.0012292
- Schuchert, C. (1905) "The mounted skeleton of Triceratops prorsus in the U.S. National Museum" American Journal of Science, Series 4, Series 4, 20, 458-459. doi:10.2475/ajs.s4-20.120.458. http://biodiversitylibrary.org/page/40245316
- Scannella, J.B. (2010) "Triceratops: a model organism for deciphering dinosaur heterochrony" Society of Vertebrate Paleontology Seventienth Anniversary Meeting Program and Abstracts Book, 158A. [abstrakt]
- Scannella, J.B. (2020) "A chasmosaurine ceratopsid premaxilla from the basal sandstone of the Hell Creek Formation, Montana" Vertebrate Anatomy Morphology Palaeontology, 8: 154-169. doi:10.18435/vamp29366
- Scannella , J.B. & Fowler, D.W. (2009) "Anagenesis in Triceratops: evidence from a newly resolved stratigraphic framework for the Hell Creek Formation" North American Paleontological Convention, abstracts volume, 148. [abstrakt]
- Scannella, J.B. & Fowler, D.W. (2014) "A stratigraphic survey of Triceratops localities in the Hell Creek Formation, northeastern Montana (2006-2010)" [w:] Wilson, G.P., Clemens, W.A., Horner, J.R. & Hartman, J.H. (red.) "Through the End of the Cretaceous in the Type Locality of the Hell Creek Formation in Montana and Adjacent Areas" The Geological Society of America Special Paper, 503, 313-332. doi:10.1130/2014.2503(12)
- Scannella, J.B. & Horner, J.R. (2010) "Torosaurus Marsh, 1891, is Triceratops Marsh, 1889 (Ceratopsidae: Chasmosaurinae): synonymy through ontogeny" Journal of Vertebrate Paleontology, 30(4), 1157-1168. doi:10.1080/02724634.2010.483632
- Scannella, J.B. & Horner, J.R. (2011) "‘Nedoceratops’: An Example of a Transitional Morphology" PLoS ONE, 6(12), e28705. doi:10.1371/journal.pone.0028705
- Scannella, J.B. & Horner, J.R. (2018) "Torosaurus or Triceratops? Assessing the postorbital horn core microstructure of the holotype of Torosaurus gladius reveals new insights into ceratopsid horn growth" Society Of Vertebrate Paleontology 78th Annual Meeting, Meeting Program & Abstracts, 210. [abstrakt]
- Scannella, J.B., Fowler, D.W., Goodwin, M.G. & Horner, J.R. (2012) "Transitional Triceratops: Details of an ontogenetic sequence from the upper Middle Unit of the Hell Creek Formation, Montana" Journal of Vertebrate Paleontology, 32(5, suplement), 166. [abstrakt]
- Scannella, J.B., Fowler, D.W., Goodwin, M.G. & Horner, J.R. (2013) "The clandestine role of heterochrony in ceratopsian evolution as revealed by juvenile Triceratops" Journal of Vertebrate Paleontology, 32(5, suplement), 206 [abstrakt].
- Scannella, J.B., Fowler, D.W., Goodwin, M.B. & Horner, J.R. (2014) "Evolutionary trends in Triceratops from the Hell Creek Formation, Montana" Proceedings of the National Academy of Sciences, 111(28), 10245-10250. doi:10.1073/pnas.1313334111
- Schlaikjer, E.M. (1935) "Contributions to the stratigraphy and palaeontology of the Goshen Hole area, Wyoming. II. The Torrington Member of the Lance Formation and a study of a new Triceratops" Bulletin of the Museum of Comparative Zoology at Harvard College, 76, 31-68. http://biodiversitylibrary.org/page/28870797
- Schuyf, P. (1969) "De kop van Triceratops cf. brevicornis HATCHER in het Mineralogisch-Geologisch Museum van de Technische Hogeschool te Delft" Grondboor & Hamer, 23(2), 67-73.
- SDSM (online 2017) http://www.sdsmt.edu/Academics/Museum-of-Geology/About/History/
- Seebacher, F. (2001) "A new method to calculate allometric length-mass relationships of dinosaurs" Journal of Vertebrate Paleontology, 21(1), 51-60. doi:10.1671/0272-4634(2001)021[0051:ANMTCA]2.0.CO;2
- Shay, K. (online 2008) http://www.joshshalek.com/?p=203
- Shetland Museum and Archives (online) https://www.shetlandmuseumandarchives.org.uk/exhibitions/smaa-archive#&gid=1&pid=10
- Shore, D. (online 2015) https://www.facebook.com/photo.php?fbid=923353404386647&set=p.923353404386647&type=3&theater
- SMF (online 2011) http://www.senckenberg.de/root/index.php?page_id=5206&PHPSESSID=biee30bi4l05khse5m6rvtvarha5oc96&year=2011&kid=1&id=1958
- Smith, M. (online 2013) http://edition.cnn.com/2013/06/03/us/triceratops-found/index.html
- Sternberg, C.M. (1949) "The Edmonton fauna and description of a new Triceratops from the Upper Edmonton member; phylogeny of the Ceratopsidae" National Museum of Canada Bulletin, 113, 33-46.
- Stock, T. (online 2015a) http://nmpdn.blogspot.com/2015/07/a-new-begining.html
- Stock, T. (online 2015b) http://nmpdn.blogspot.com/2015/07/harleys-head-and-his-leaf-munching.html
- Sues, H.-D. & Galton, P.M. (1987) "Anatomy and classification of the North American Pachycephalosauria (Dinosauria: Ornithischia)" Palaeontographica Abteilung A, 198(1-3), 1-40.
- Sullivan, R.M. & Lucas, S.G. (2010) "A New Chasmosaurine (Ceratopsidae, Dinosauria) from the Upper Cretaceous Ojo Alamo Formation (Naashoibito Member), San Juan Basin, New Mexico" [w:] Ryan, M.J., Chinnery, B.J. & Eberth, D.A. (red.) "New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium" wyd. Indiana University, Bloomington, 169-178.
- Swanson, A. (online 2013) http://csceagle.com/2013/02/13/geology-professors-digs-yield-more-than-awards/
- Swiderski, M. (online 2008) http://www.flickr.com/86426643@N00/2342505208
- Switek, B. (online 2011): http://blogs.smithsonianmag.com/dinosaur/2011/01/the-great-triceratops-debate-continues/
- Switek, B. (online 2013) http://phenomena.nationalgeographic.com/2013/06/11/chipping-away-at-triceratops-hype/
- Tanke, D.H. (online 2014) https://www.facebook.com/groups/7636841670/permalink/10151950262541671/
- Tanke, D. (online 2016) https://www.facebook.com/groups/7636841670/permalink/10153707571891671/
- Tanke, D. (online 2018) https://www.facebook.com/groups/7636841670/posts/10155343949206671/
- Tanke, D.H. & Farke, A.A. (2006) "Bone resorption, bone lesions, and extracranial fenestrae in ceratopsid dinosaurs: a preliminary assessment" [w:] Carpenter, K. [red.] "Horns and Beaks: Ceratopsian and Ornithopod Dinosaurs" Indiana University Press, Bloomington, 319-347.
- The Boomtown (online 2013) https://www.facebook.com/photo.php?fbid=623654997663992
- Thompson, S. & Holmes, R. (2007) "Forelimb stance and step cycle in Chasmosaurus irvinensis (Dinosauria: Neoceratopsia)" Palaeontologia Electronica, 10(1), 5A.
- Timmons, J. (online 2014) http://mostlymammoths.wordpress.com/2014/12/20/the-undetermined-fate-of-a-rare-triceratops-fossil/
- TMP (online) https://hermis.alberta.ca/TMP/Details.aspx?ObjectID=TMP1984.039.0012&dv=True
- Tokaryk, T.T. (1986) "Ceratopsian dinosaurs from the Frenchman Formation (Upper Cretaceous) of Saskatchewan" Canadian Field Naturalist, 100(2), 192-196.
- Tokaryk, T.T. (2002) A Humerus from a Juvenile Triceratops sp. (Ornithischia; Ceratopsidea) from the Frenchman Formation (Late Maastrichtian), Saskatchewan Summary of Investigations 2002(1), Saskatchewan Geological Survey; Miscellaneous Report, 2004-4.1, 151-154.
- Tokaryk, T.T. (2009) "Head-hunting in Saskatchewan: the history of Triceratops" [w:] Tokaryk, T., Snively, E. & Bryant, H. (red.) "Frenchman Formation Terrestrial Ecosystem Conference, Program, Abstracts of Papers, Workshops and Field Trip" Royal Saskatchewan Museum Contribution to Science, 12, 61-63.
- Tsuihiji, T. (2010) "Reconstructions of the axial muscle insertions in the occipital region of dinosaurs: evaluations of past hypotheses on Marginocephalia and Tyrannosauridae using the Extant Phylogenetic Bracket approach" The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology, 293(8), 1360–1386. doi:10.1002/ar.21191
- UCMP (online) - University of California Museum of Paleontology collections http://ucmpdb.berkeley.edu
- Ukrainsky, A. S. (2007) "A new replacement name for Diceratops Lull, 1905 (Reptilia: Ornithischia: Ceratopsidae)" Zoosystematica Rossica, 16(2), 292.
- USNM (online 2014) http://nmnh.typepad.com/smithsonian_fossils/2014/01/triceratops-storage-jacket.html
- USNM (online) http://collections.si.edu/search/index.htm
- WBRZ (online) http://www.wbrz.com/videos/meet-jason-the-triceratops/
- Weishampel, D.B., Barrett, P.M., Coria, R.A., Le Loeuff, J. Xu, X., Zhao, X., Sahni, A., Gomani, E.M.P. & Noto, C.R. (2004) "Dinosaur Distribution" [w:] Weishampel, D.B., Dodson, P. & Osmólska, H. "The Dinosauria" University of California Press, 517-606.
- Wilson, G.P. (2005) "Mammalian faunal dynamics during the last 1.8 million years of the Cretaceous in Garfield County, Montana" Journal of Mammalian Evolution, 12, 53-76. doi:10.1007/s10914-005-6943-4
- Wilson, G.P. (2014) "Mammalian extinction, survival, and recovery dynamics across the Cretaceous-Paleogene boundary in northeastern Montana, USA" [w:] Wilson, G.P., Clemens, W.A., Horner, J.R. & Hartman, J.H. (red.) "Through the End of the Cretaceous in the Type Locality of the Hell Creek Formation in Montana and Adjacent Areas" The Geological Society of America Special Paper, 503, 365-392. doi:10.1130/2014.2503(15)
- Wilson, J.P. & Fowler, D.W. (2017) "First confirmed identification of juvenile Triceratops epiparietals" Cretaceous Research, 70, 71-76. doi:10.1016/j.cretres.2016.10.005
- Wick, S.L. & Lehman, T.M. (2013) "A new ceratopsian dinosaur from the Javelina Formation (Maastrichtian) of West Texas and implications for chasmosaurine phylogeny" Naturwissenschaften, 100(7), 667-682. doi:10.1007/s00114-013-1063-0
- "WitmerLab" (online) http://www.oucom.ohiou.edu/dbms-witmer/dinoskulls04.htm
- Witton, M. (online 2017) http://markwitton-com.blogspot.co.uk/2017/11/can-we-predict-horn-shapes-of-fossil.html
- Wolfe, D.G. & Kirkland, J.I. (1998) "Zuniceratops christopheri n. gen. & n. sp., a ceratopsian dinosaur from the Moreno Hill Formation (Cretaceous, Turonian) of west-central New Mexico" Lower and Middle Cretaceous Terrestrial Ecosystems, New Mexico Museum of Natural History and Science Bulletin, 24, 307-317.
- Wolff, E.D.S. (2007) "Oral pathology of the Archosauria: bony abnormalities and phylogenetic inference" dysertacja doktorska, Montana State University, Bozeman, 1-312. http://scholarworks.montana.edu/xmlui/handle/1/2570
- Woollatt, B. (online) http://photosynth.net/view.aspx?cid=368e66d6-0f62-4e00-a601-f4f5e74f0ed1
- Woollatt, B. (online 2016) https://www.facebook.com/photo.php?fbid=10153351159451962&set=pcb.10153356455691671&type=3
- Wu, X-c., Brinkman, D.B., Eberth, D.A. & Braman, D.R. (2007) "A new ceratopsid dinosaur (Ornithischia) from the uppermost Horseshoe Canyon Formation (upper Maastrichtian), Alberta, Canada" Canadian Journal of Earth Sciences, 44(9), 1243-1265. doi:10.1139/e07-011
- You, H. & Dodson, P. (2004) "Basal Ceratopsia" [w:] Weishampel, D.B., Dodson, P. & Osmólska, H. "The Dinosauria" University of California Press, 478-493.
- YPM (online 2013) http://www.youtube.com/watch?v=XTVpo3grXMY
- YPM (online) (katalog, Peabody Museum of Natural History, Division of Vertebrate Paleontology, Yale University) http://peabody.yale.edu/collections
{kind=link}
a.jpg){kind=link}
{kind=link}
- ↑ 5 t za Seebacher (2001) - na podstawie opartej na BSP 1964 I 458 i USNM 4842 rekonstrukcji z Ostrom i Wellnhofer (1986), z długością ciała 8 m. 13,5 t za Benson i in. (2014): AMNH 5033 (przynależność taksonomiczna niepewna - szkielet pozaczaszkowy, który jest zmontowany z kilkoma innymi z AMNH [5116, 5045, 5039; odlewy: 970, 971, 973] w okaz o długości 7,1 m [Osborn, 1933]); USNM 4842 - 11 t. Paul (1997) podał 6-7 t, zaznaczył jednak, że istnieją okazy, których masa sięgała 10 ton a odciski stóp sugerują 12-19 t. Wybrane inne oszacowania (chronologicznie): Alexander (1985): USNM 4842 - 6,1/9,4 t. Alexander (1989) - 6 t (za Seebacher, 2001). Farke (online 2005): TCM 2001.93.1 - 5,2 t; USNM 4842 - 10-11 t. Paul (2010) - 9 t. Dane Paula (online 2010), do których odsyła książka, to: USNM 4276 (przynależność taksonomiczna niepewna) - 9,3 t; USNM 4842 - 6,4 t (tą samą wartość dla tego okazu podał Longrich [2011], nie wskazał jednak źródła tego twierdzenia); MWC 7584 (podano jedynie oznaczenie BYU) - 9 t. Longrich (2011): LACM 150076 (przynależność taksonomiczna niepewna) - 7,3 t. Campione i Evans (2012): NSM PV 20379 - 7,4 t (5,3 t metodą Andersona i in., 1985). Benson i in. (2017): NSM PV 20379 – 6,1 t; HMNS PV.1506 – 11,4 t, USNM 4842 – 11,4 t. Henderson (1999) posłużył się modelem długości ok. 6,2 m (oszacowane za ryc. 10D), którego oszacowana masa to 3,9 t, zaś Colbert (1962) rzeźbą w skali 1/15 lub 1/16 i uzyskał 8,5 t. Najczęściej szacowany okaz - USNM 4842: ok. 6 m długości i 6-11 t - składa się z kilku osobników różnej wielkości (Schuchert, 1905; Gilmore, 1906). Czaszka, którą zamieszczono w szkielecie (USNM 2100), ma ok. 183 cm (6 stóp) (Gilmore, 1906) i pochodzi od mniejszego osobnika (Ostrom i Wellnhofer, 1986, s. 132) a kość udowa mierzy 113,7 cm (Chinnery, 2004).
- ↑ Określenie, kiedy żył Triceratops jest problematyczne, głównie dlatego, że dostępne dane opierają się na umiejscowieniu granicy kreda-paleogen 65,5 Ma a obecnie granicę przesuwa się na 66 Ma (International Chronostratigraphic Chart, International Commission on Stratigraphy, 2013). Wiek Eotriceratops, wcześniejszego i bardziej bazalnego, prawdopodobnego przodka Triceratops, określało się na 68-67,6 Ma (Wu i in., 2007; Eberth i in., 2013), zaś Triceratops na 66-65,5 (Arbour i in. 2009, ryc. 7) lub 67-65,5 (Sampson i Loewen, 2010, s. 410). Eberth i in. (2013) wskazali wiek ogniwa Carbon, z którego pochodzi Eotriceratops, na 68,4-67,5 Ma, Fowler (2017) zamieścił Eotriceratops w przedziale 68,3-68,2 Ma a Eberth i Kamo (2020) wydatowali go na 68,8 ± 0,1 Ma. Formacja Hell Creek zawiera osady dokumentujące prawdopodobnie 1-2 miliony lat (Scannella i in., 2014): 1,05-1,9 mln lat (prawdopodobnie ok. 1,36 mln), więc 66,8-65,5 Ma (Hicks i in., 2002), 67,7-65 Ma (Lund i in., 2002) a według nowszych oszacowań 2,1 mln lat – 67,7-65,58 Ma (Wilson, 2005), 1,8-2 mln lat – 67,5/67,3-65,5 Ma (Wilson, 2014, s. 371). Formacja Hell Creek nie jest więc starsza niż 68,2 Ma (dolna granica C30n - Ogg, 2012, s. 92, por. ryc. 11 w Hicks i in., 2002). Fowler (2017) datuje część zawierającą Triceratops i Torosaurus (określony przez niego jako Triceratops sp. 1) formacji Hell Creek w Montanie na niecały 66,8-66 Ma a zawierającą Triceratops 66,5-66 Ma. Dokładne datowanie formacji Lance jest nieznane (Fowler, online 2016; 2017).
- ↑ http://www.filmweb.pl/film/Zaginiony+świat-1925-89112
- ↑ http://en.wikipedia.org/wiki/The_Lost_World_(1925_film)#Prehistoric_Creatures
- ↑ http://www.youtube.com/watch?v=55W_XCWckpI
- ↑ http://www.youtube.com/watch?v=Msbt66Z2sjY
- ↑ http://www.filmweb.pl/film/Milion+lat+przed+naszą+erą-1966-32345
- ↑ http://www.youtube.com/watch?v=Tx7vCJoCszk
- ↑ http://www.filmweb.pl/film/Last+Dinosaur%2C+The-1977-232483
- ↑ http://www.youtube.com/watch?v=B619J7VeYAY
- ↑ http://www.youtube.com/watch?v=RF6W9m1Malk
- ↑ http://www.youtube.com/watch?v=_hQm9sLhdbM
- ↑ http://www.filmweb.pl/serial/Zaginiony+ląd+-+Dolina+Jurajska-1991-106789
- ↑ http://www.youtube.com/watch?v=dmcx0PWPYj0
- ↑ http://www.filmweb.pl/film/Pradawny+Ląd+1%3A+Pierwsza,+wielka+przygoda-1988-10533
- ↑ http://www.filmweb.pl/serial/Dino-Riders-1988-225737
- ↑ http://www.youtube.com/watch?v=tpuhLkh358Y
- ↑ http://www.filmweb.pl/film/Dink%2C+mały+dinozaur-1989-247563
- ↑ http://www.youtube.com/watch?v=Nv4_TUx2Ho0
- ↑ http://www.filmweb.pl/serial/Mała+syrenka-1992-96111
- ↑ http://www.filmweb.pl/serial/Niebezpieczne+dinozaury-1997-374982
- ↑ http://www.youtube.com/watch?v=SqgJ6wzSW8k&feature=fvw
- ↑ http://blabbermouth.net/news.aspx?mode=Article&newsitemID=187816 (kopia w https://archive.is/B1CBo#selection-879.167-879.176 )
- ↑ http://en.wikipedia.org/wiki/Triceratops_(band)
- ↑ http://brickset.com/detail/?Set=5955-1
- ↑ http://brickset.com/detail/?Set=5975-1
- ↑ http://brickset.com/detail/?Set=4892-1
- ↑ http://brickset.com/detail/?Set=7604-1
- ↑ http://brickset.com/detail/?Set=5885-1
- ↑ https://tyranttr.deviantart.com/art/Triceratops-Prorsus-Saurian-705540983
- ↑ https://www.deviantart.com/arvalis/art/Saurian-Triceratops-Lifecycle-589354279
Skróty instytucjonalne
AMNH, American Museum of Natural History, Nowy York; ASU, Appalachian State University, Boone, Karolina Północna; BCGM, Bob Campbell Geology Museum, Clemson, Karolina Południowa; BHI, Black Hills Institute of Geologcial Research, Hill City; BMRP, Burpee Museum of Natural History, Rockford; BSM, Birmingham Science Museum, Birmingham; BSP, Bayerische Staatsamlung fur Palaontologie und Historische Geologie, Monachium; BYU, Brigham Young University Earth Science Museum, Provo, Utah; CCM, Carter County Museum, Ekalaka; CM, Carnegie Museum of Natural History, Pittsburgh; CNM, Canadian Museum of Nature, Ottawa; CSC, Chadron State College, Chadron, Nebraska (Eleanor Barbour Cook Museum); DMC, Dickinson Museum Center / Badlands Dinosaur Museum, Dickinson, Dakota Północna (wcześniej Dakota Dinosaur Museum); DMNH, Denver Museum of Nature and Science, Denver (wcześniej Denver Museum of Natural History); EM, Eastend Historical Museum, Eastend; ESU, Emporia State University, Emporia, Kansas; FMNH, Field Museum of Natural History, Chicago; FPDM, Fukui Prefectural Dinosaur Museum, Fukui; GCM, Garfield County Museum, Jordan, Montana; GLAHM, Hunterian Museum and Art Gallery, Glasgow; GMNH, Gunma Museum of Nature and Science, Tokio; GP, Glenrock Paleontological Museum, Glenrock, Wyoming; GRM, Grand River Museum, Lemmon, Dakota Południowa; HMNS, Houston Museum of Natural Science, Houston; KUVP, University of Kansas Natural History Museum, Lawrence; LACM, Natural History Museum of Los Angeles County, Los Angeles; LASM, Louisiana Art & Science Museum, Baton Rouge, Louisiana; MCZ, Museum of Comparative Zoology, Harvard University, Cambridge; MGM, Mineralogisch-Geologisch Museum, Technische Universiteit Delft, Delft; MNHN, Muséum national d’Histoire naturelle, Paryż; MOR, Museum of the Rockies, Bozeman; MOS - Museum of Science, Boston; MPM, Milwaukee Public Museum, Milwaukee; MRF, Marmarth Research Foundation, Marmarth, Dakota Północna; MSP, Makoshika State Park, Glendive, Montana; MWC, Dinosaur Journey Museum, Museum of Western Colorado, Fruita; NBC, Naturalis Biodiversity Center, Leiden, Holandia; NDGS, North Dakota Geological Survey, Bismarck, Dakota Północna; NDHC, North Dakota Heritage Center, Bismarck, Dakota Północna; NSM, National Museum of Nature and Science, Tokio; OMNH, Oklahoma Museum of Natural History, Norman; PTRM, Pioneer Trails Regional Museum, Bowman, Dakota Północna; ROM, Royal Ontario Museum, Toronto; RSM, Royal Saskatchewan Museum, Regina, Saskatchewan; TMP, Royal Tyrrell Museum of Palaeontology, Drumheller; SDSM, South Dakota School of Mines, Rapid City; SMF, Senckenberg Naturmuseum, Frankfurt nad Menem; SMM, Science Museum of Minnesota, St. Paul; SMNH, Royal Saskatchewan Museum, Regina (wcześniej Saskatchewan Museum of Natural History); SUP, Shenandoad University, Winchester; TCM, The Children's Museum, Indianapolis; TLAM, Timber Lake Area Museum, South Dakota; UCM, University of Colorado, Boulder, Colorado; UCMP, University of California Museum of Paleontology, Berkeley; UCM, University of Colorado Museum, Boulder; UM, University of Michigan Museum of Paleontology, Ann Arbor; UND, Paleontology Collection, University of North Dakota, Grand Forks; USNM, United States National Museum of Natural History, Waszyngton; UWGM, University of Wisconsin-Madison Geology Museum, Madison; WDC, Wyoming Dinosaur Center, Thermopolis; WCCH, Weld County Court House, Greeley, Colorado; WFQ, Warfield Fossil Quarries; WMUC, Walker Musuem of the University of Chicago, Chicago; YPM, Yale Peabody Museum, New Haven;